Patrones de supervivencia de Pinus cembroides Zucc. en una comunidad montañosa del NE de México
Jaime F. García*
CIENCIA UANL / AÑO 23, No.99 enero-febrero 2020
DOI: https://doi.org/10.29105/cienciauanl23.99-1
RESUMEN
Se espera que los efectos del calentamiento global incrementen fenómenos meteorológicos extremos como sequías y lluvias torrenciales, estableciendo la necesidad de integrar sus consecuencias al manejo de ecosistemas. La supervivencia de plántulas de Pinus cembroides en una comunidad montañosa del noreste de México, debido a: (1) la pendiente del suelo, (2) las plantas nodrizas, (3) el hábitat y (4) la variación interanual en lluvias, fue examinada y se probaron hipótesis para identificar patrones de supervivencia de plántulas. Los resultados indican: (i) mayor supervivencia de plántulas en sitios con pendientes >30°, (ii) la asociación nodriza-protegida incrementó la supervivencia de plantas, (iii) entre hábitats la supervivencia fue mayor en vegetación densa y (iv) mayor supervivencia de plántula en vegetación densa en el año húmedo. Los manejadores deberán considerar acciones que promuevan el reclutamiento de plantas a través de la conservación de sitios ideales para el establecimiento de plantas.
Palabras clave: supervivencia, manejo, montaña, vegetación densa.
ABSTRACT
The effects of global warming are expected to increase extreme weather events such as droughts and torrential rains, establishing the need to integrate its consequences to the ecosystem management. Survival of Pinus cembroides seedlings in a mountainous community of Northeast Mexico, due to: (1) slope of the soil, (2) nurse plants, (3) habitats and (4) annual variation in rainfall, was examined and hypotheses were tested to identify seedling survival patterns. The results indicate: (i) there is greater seedling survival at sites with slopes> 30 °, (ii) the nurse-protected association increased plant survival, (iii) between habitats, survival was higher in dense vegetation and (iv) there is greater seedling survival in dense vegetation in the wet year. Managers should consider actions that promote the plants recruitment, through the conservation of sites ideal to plants establishment.
Keywords: survival, management, mountain, dense vegetation.
Evidencias del cambio climático han sido documentadas por investigadores en términos de cambios en temperaturas (Hughes, 2000). Lo que está alterando la distribución de plantas (McLachlan et al., 2005) y animales (Peterson et al., 2002). El impacto del cambio climático sobre las plantas, en sus ecosistemas naturales, ha sido documentado en la investigación ecológica actual (Müller, 2017). Las proyecciones climáticas predicen alteraciones en la frecuencia e intensidad de fenómenos meteorológicos extremos, como sequías y lluvias torrenciales (Gobiet et al., 2014; IPCC, 2013). Las sequías extremas han sido asociadas a la reducción en la absorción de CO2 y a menor crecimiento de plantas, reduciendo el potencial de sumidero de carbono del ecosistema (Frank et al., 2015).
México es un país altamente vulnerable a las sequías, 52% de su territorio está clasificado como árido o semiárido (Esparza, 2014), con escasas lluvias a lo largo del año, las sequías se presentan de manera frecuente. En la zona montañosa del sur de Nuevo León, Pinus cembroides (piñón piñonero) es la especie de plantas arbóreas más ampliamente distribuida en el ecosistema y sus semillas son colectadas para consumo humano (García y Jurado, 2015). En esta región, históricamente se presentan índices pluviales bajos que oscilan entre 279.4 y 517.1 mm (Conagua, 2009), por lo que las sequías son recurrentes. No obstante, en ecosistemas semiáridos como el de la región montañosa del sur de Nuevo León, la variación interanual en precipitaciones puede jugar un papel crucial en la dinámica de las poblaciones vegetales. En esta región montañosa los efectos de la sequía pueden no estar generalizados en el ecosistema, debidoa la variación en la topografía local y a las múltiples interacciones entre plantas que pueden reducir o incrementar la supervivencia de plantas. Un aumento en la humedad aprovechable del suelo causada por lluvias torrenciales puede incrementar el establecimiento de plantas (García, 2011), y limitarlo en años secos.
El presente estudio examina factores asociados a la supervivencia de plántulas en el estadio temprano de establecimiento, lo que permitirá identificar patrones de supervivencia y contribuirá a predecir futuras distribuciones de la especie a escala regional. Se examina el efecto de la pendiente del suelo y de las plantas nodrizas en plántulas ≤ 20 cm, las cuales representan a los individuos de la regeneración, se estudia la biología de establecimiento de plántulas sembradas y plantadas, lo que permitirá: i ) cuantificar el efecto de la pendiente del suelo en la supervivencia de plántulas ≤ 20cm, ii) determinar si la supervivencia de plántulas ≤ 20 cm es mayor bajo el dosel de la vegetación, iii) cuantificar el efecto del hábitat en la supervivencia de plántulas y iv) establecer el efecto de la variación interanual en precipitaciones en la supervivencia de plántulas.
Se probaron cuatro principales hipótesis: (1) la supervivencia de plántulas en suelos con diferentes grados de pendiente varía a lo largo del ecosistema debido a hábitats más o menos estresantes, (2) plantas nodrizas del sotobosque son mecanismos de facilitación planta-planta y promueven mayor supervivencia de plantas, (3) la supervivencia de plántulas es más alta en hábitats con mayor cubierta vegetal y (4) la variación interanual en precipitaciones incrementa la supervivencia de plántulas en el hábitat ideal en año húmedo.
MÉTODOS
Sitio de estudio
El estudio se realizó desde el otoño de 2007 al de 2014, en cinco comunidades montañosas ubicadas en La Ascensión, Aramberri, Nuevo León, cuyas coordenadas geográficas son: I) 24º 30’ 43 N, 99º 36’ 34 W; II) 24º 31’ 23.3’’ N, 99º 59’ 28.9’’ W; III) 24º 28’ 39.6’’ N, 99º 59’ 34.2’’ W; IV) 24º 33’ 52.8’’ N, 99º 37’ 50.3’’W, y (V) 24º 30’ 55.2’’ N, 99º 53’ 59.3’’W. El clima es fresco, medio seco con una precipitación anual que en el periodo de estudio osciló entre 374.1 a 1007.4 mm (figura 1a-b). Se presentan escasas lluvias en primavera y ligeramente intensas en verano y principios del otoño. Las temperaturas promedio fueron: media anual 16.8°C, máxima 24.5°C y mínima de 8.7°C. En invierno frecuentemente se presentan temperaturas que oscilan entre -1 y -6°C (Comisión Nacional del Agua, estación La Ascensión).
EFECTO DE LA PENDIENTE DEL SUELO
Para establecer el efecto de la pendiente en la supervivencia de plántulas de P. cembroides ≤ 20 cm, en diciembre de 2012 se seleccionaron cuatro áreas con dos tipos de pendiente, en cada área se establecieron cuatro sitios y en cada sitio se contaron los individuos en dos parcelas de 10×25 m y todos los individuos fueron registrados en un mapa. Las pendientes se clasificaron en: a) baja a moderada 0-30° y b) pronunciada >30°. En diciembre de 2014 las plántulas registradas dentro de cada parcela se contaron e identificaron como vivas o muertas, y se compararon los niveles de supervivencia entre pendientes con una prueba t de Student para muestras independientes.
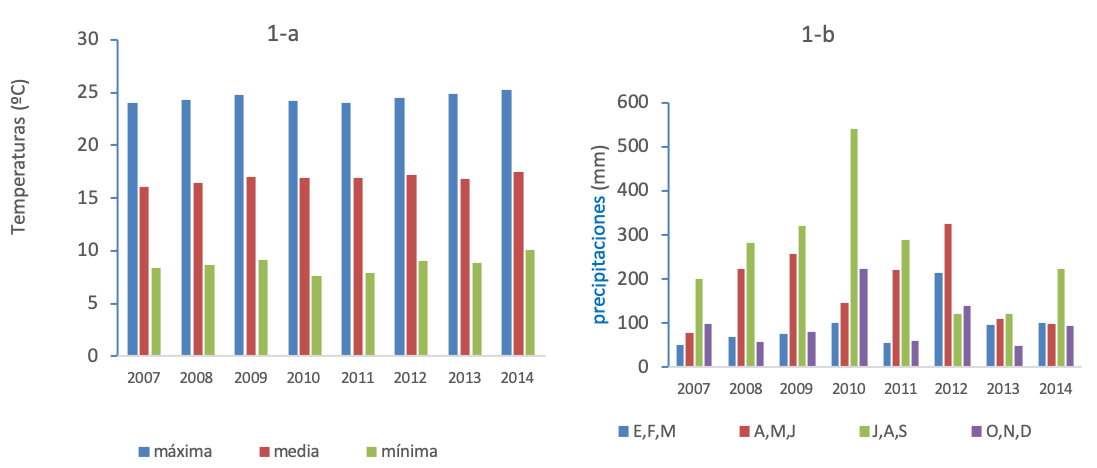
Figura 1. Registro de temperaturas (1-a) y precipitaciones (1-b) en la zona de estudio (datos proporcionados por la CNA).
Efecto de las plantas nodrizas
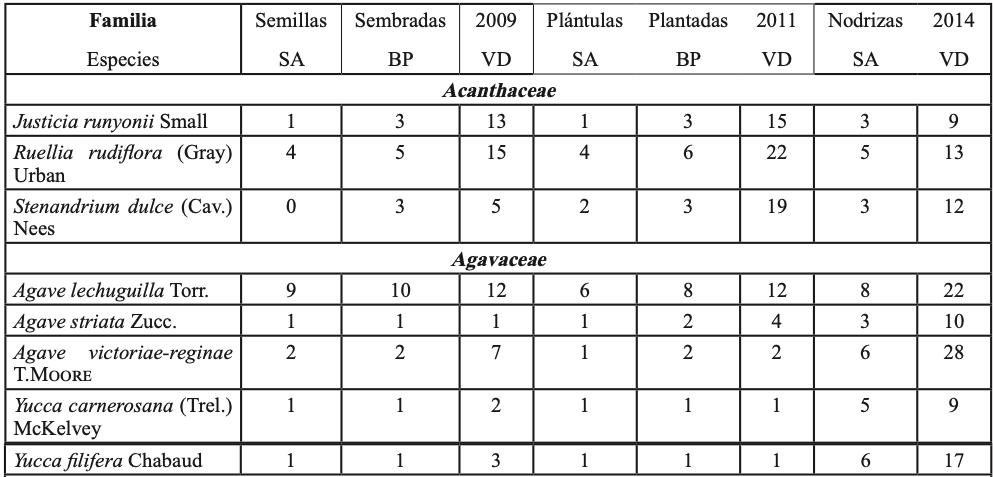
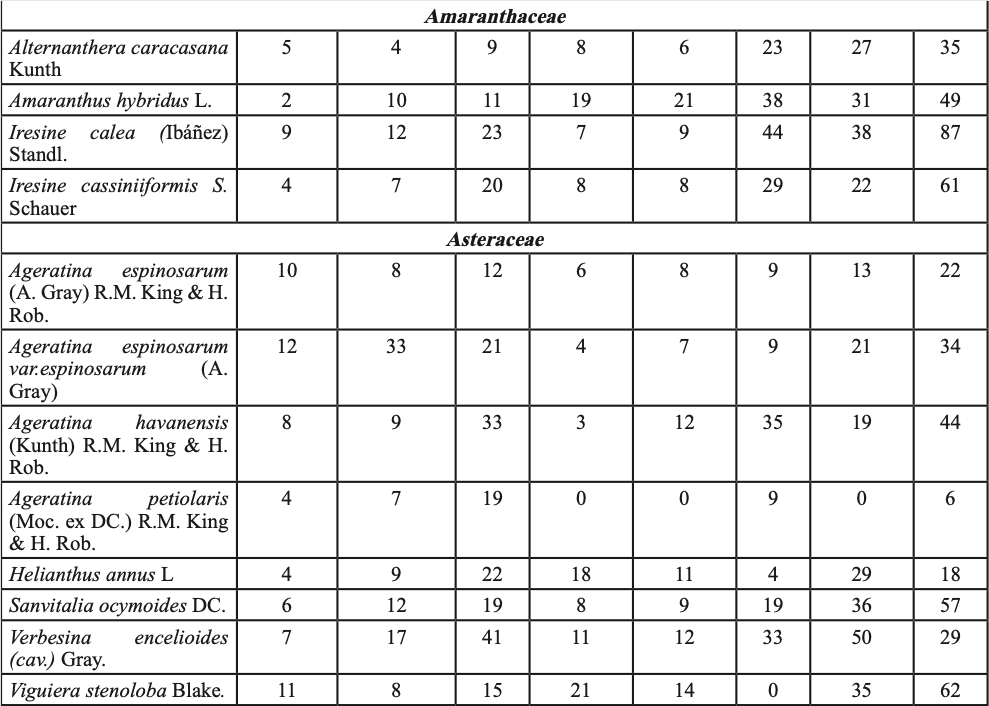
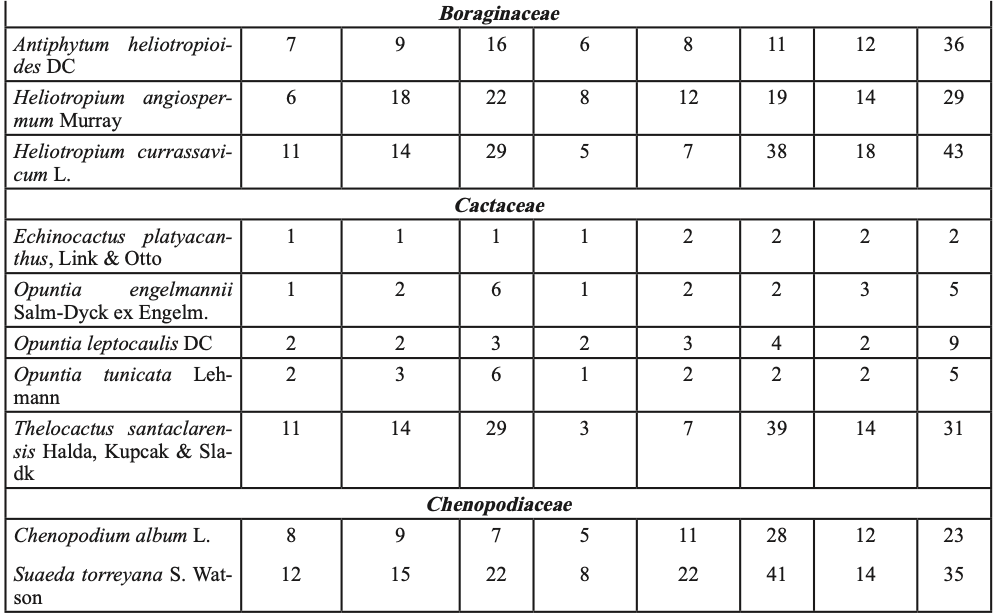
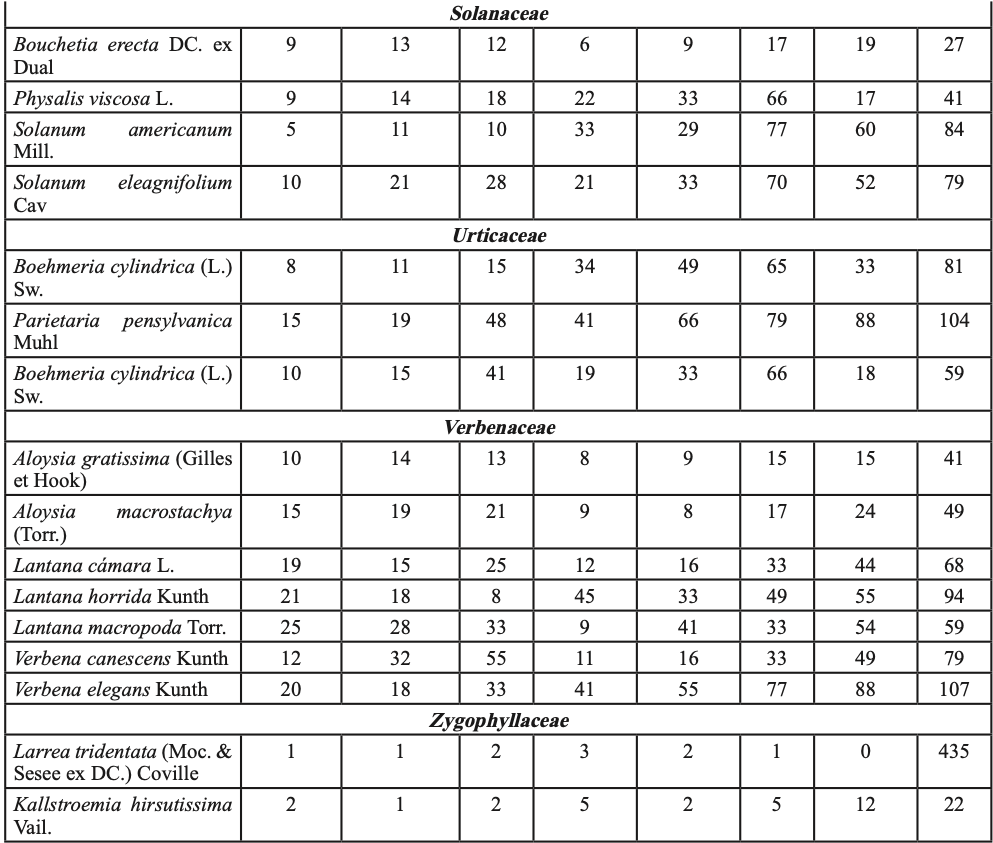
Para probar la hipótesis de que las plántulas de P. cembroides ≤ 20 cm creciendo con plantas del sotobosque presentan mayor tasa de supervivencia, en el invierno de 2012 (un año con abundantes lluvias de primavera, figura 1-b), en las cinco áreas de muestreo, se establecieron cuatro sitios por área y en cada sitio se muestrearon los individuos en tres parcelas de 10×25 m, se ubicaron las plántulas de P. cembroides ≤ 20cm y se clasificaron creciendo en asociación con: (i) vegetación del sotobosque (hierbas, pastos, arbustos y cactus, tabla I), (ii) bajo el dosel de P. cembroides y (iii) sobre suelo despejado con baja cobertura de vegetación. En el invierno de 2014 las plántulas fueron contadas y clasificadas como vivas o muertas y los niveles de supervivencia entre los tres hábitats: (i) vegetación del sotobosque, (ii) bajo el dosel de P. cembroides y (iii) suelo con escasa cobertura de vegetación, fueron comparados utilizando la prueba de Kruskal-Wallis para K muestras independientes.
Efecto del hábitat en la supervivencia de plántulas
Para probar el efecto del hábitat en la supervivencia de P. cembroides se colectaron semillas del piñonero durante el otoño de 2007. La colecta fue realizada de poblaciones naturales de P. cembroides distribuidas en los sitios de muestreo. En cada sitio se seleccionaron 50 árboles con el fin de abarcar la variación genética natural de las poblaciones, la distancia entre árboles fue de 50 m. Se seleccionaron árboles fenotípicamente superiores y se colectaron semillas sin evidencias de parasitismo o daños por insectos (García-Pérez et al., 2007). Las semillas fueron mezcladas y puestas a secar en papel estraza con bolsas de gel de sílice para controlar la humedad y permitir su maduración; antes de cada ensayo las semillas fueron escarificadas con papel lija #40, tratamiento reportado como eficaz para causar la germinación de semillas con integumentos duros en el noreste de México (Jurado et al., 2006).
En septiembre de 2008 y agosto de 2009, época del año que coincide con la germinación y el establecimiento de especies nativas en la región (García y Jurado, 2015), fueron elegidos tres sitios con hábitats contrastantes: i) suelo abierto (SA) con escasa vegetación preexistente, ii) bajo dosel de pino (BP) a un radio de 2 m de la base del pino y iii) vegetación densa (VD) o de sotobosque (tabla I). En cada hábitat se escogieron treinta parcelas de 2×2 m (15/año). El número de parcelas utilizado fue definido de acuerdo a la fórmula desarrollada por Lozano-Rivas (2011).
Se sembraron 40 semillas por parcela a una distancia entre semillas de 40 cm. Las semillas para su emergencia y crecimiento dependieron de las condiciones ambientales. El ensayo fue replicado en agosto de 2009 y la supervivencia de plantas fue reportada cada semana por 18 meses. La supervivencia acumulada de plantas fue calculada como porcentaje de supervivencia de plántulas = (número de plántulas vivas/número de plántulas emergidas)*100. La supervivencia de plantas de semillas sembradas entre hábitats (SA, BP y VD) fue evaluado mediante el uso de ANOVA (α = 0.05) de un factor, se utilizaron pruebas múltiples de comparación de promedios de Tukey cuando los ANOVA indicaron diferencias estadísticas significativas. E intervalos de confianza (α =0.05) en la gráfica para resaltar las diferencias de supervivencia entre hábitats. Previo a los análisis, los datos fueron transformados mediante la función arcoseno para ajustarse a la distribución normal (Sokal y Rohlf, 1995).
Efecto de la variación interanual en precipitaciones en la supervivencia de plántulas
Para contrastar el efecto interanual debido a las precipitaciones de verano-otoño de 2010 y 2011, en plántulas plantadas en tres hábitats (SA, BP y VD), se preparó sustrato a base de tierra y hojarasca proveniente de los tres hábitats y se depositó en bolsas de plástico de 6128.5 cm3. En septiembre de 2009 y 2010 se sembraron semillas de P. cembroides previamente escarificadas en los contenedores (una semilla por contenedor). Las semillas fueron regadas cada tercer día, para su germinación y crecimiento. Posteriormente, las plántulas fueron plantadas en cada hábitat en septiembre de 2010 y agosto de 2011, cada planta tenía aproximadamente 15 meses de edad. Para la plantación fueron seleccionados individuos de altura uniforme para eliminar sesgo en los resultados. La altura de plantas (cm) y el diámetro del tallo (mm) en 2010 y 2011 fueron: i) en SA 22.5 ± 1.8, 13.3 ± 4.5; 26.9 ± 2.9, 15.1 ± 6.7; ii) BP 24.1 ± 2.3, 16.9 ± 6.8; 20.3 ± 1.7, 16.5 ± 9.9, y iii) en VD 23.2 ± 3.4, 15.6 ± 11.3; 26.0 ± 2.6, 16.3 ± 8.1 (̅X± DE.). Se registraron un total de 600 plántulas: (10 plántulas x 3 hábitats x 2 años x 10 sitios replicados). Para el trasplante, el cepellón fue cuidadosamente retirado de la bolsa y depositado en hoyos de 20 cm de diámetro por 30 cm de profundidad. Para evitar el estrés postransplante, las plántulas fueron regadas con aproximadamente 5 l de agua in situ cada tercer día por dos semanas. Las plántulas fueron rociadas con una dilución de insecticida (piretrina sintética) una vez por semana, para controlar el daño por insectos defoliadores de la orden lepidóptera e himenóptera, los cuales, en su fase larvalo adulta, se alimentan del follaje de la planta, ocasionando una reducción del área foliar, lo que trae como consecuencia el debilitamiento de la planta. Las plantas fueron ubicadas en un área heterogénea boscosa de 10 ha, que en los tres hábitats albergó diferentes especies de plantas (tabla I). Se tomaron datos cada semana por un total de 18 meses, y el porcentaje de supervivencia de plántulas fue calculado como (número de plántulas vivas/total de plantas)*100. El efecto interanual debido a las lluvias, en plántulas trasplantadas entre los tres hábitats (tabla I) se evaluó con ANOVA de dos factores, éstos fueron: 1. la exposición a la lluvia de verano-otoño: de (i) 2010 y (ii) 2011 y 2. hábitats: (iii) suelo abierto SA, (iv) bajo dosel de pino BP y vegetación densa VD. Se generaron seis tratamientos (2 años x 3 hábitats): 1) SA-2010, 2) BP-2010, 3) VD-2010, 4) SA-2011, 5) BP-2011 y 6) VD-2011. Se utilizaron pruebas de Tukey e intervalos de confianza (α =0.05) en las gráficas para resaltar las diferencias de supervivencia entre hábitats y años. Los datos fueron transformados mediante la función arcoseno para ajustarse a la distribución normal.
RESULTADOS
La pendiente del suelo
Se registró un total de 1208 plántulas, 609 en la pendiente baja 0-30° y 599 en la pendiente pronunciada >30°. Sobrevivieron 542 plántulas, 192 en la pendiente baja y 350 en la pendiente pronunciada, % de supervivencia = 44.9%. Los resultados (t = 10.3, g. l. = 15, p < 0.05; figura 2-a) indican diferencias en la supervivencia de plantas entre las laderas 0-30° (35.4%) y >30° (64.6%). La mayor supervivencia de plántulas se presentó en la pendiente >30°. En plantaciones forestales en suelos severamente erosionados, Gómez-Romero et al. (2012) mencionan similar tendencia en P. cembroides > a 50cm.
Plantas nodrizas
Hubo un total de 1625 plántulas, 601 en suelo abierto, 499 bajo el dosel del pino y 525 en la vegetación del sotobosque, sobrevivieron 870 individuos, % de supervivencia = 53.5%. Se registraron 173 plántulas en suelo abierto (% de supervivencia = 19.9%), 214 bajo el dosel del pino (% de supervivencia 24.6%), mientras que en el sotobosque se registraron 483 individuos (% de supervivencia = 55.5%). Los hábitats con alta cobertura de plantas se asociaron significativamente con la supervivencia (χ2 = 14.08, g. l. = 2, p < 0.02; figura 2-b). La alta supervivencia de P. cembroides bajo plantas nodrizas indica una interacción positiva con la vegetación del sotobosque (tabla I), lo que incrementa el gradiente de supervivencia.
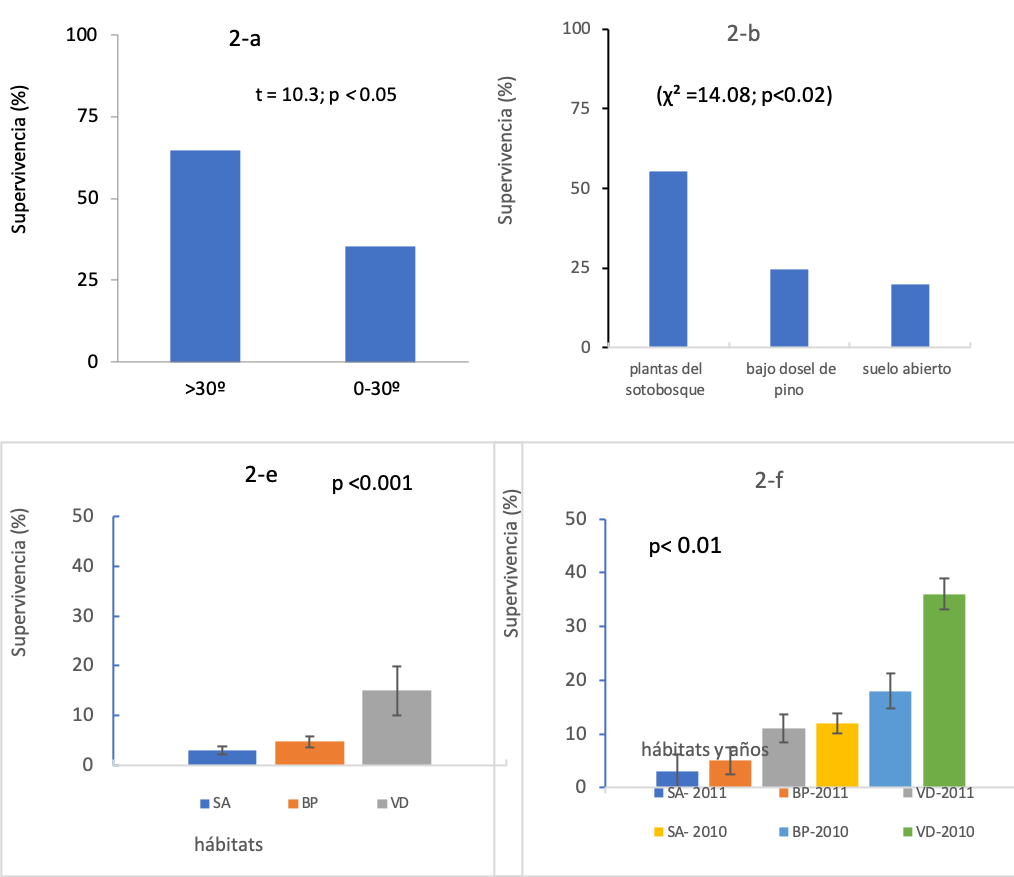
Figura 2. Supervivencia de plántulas de P. cembroides: (a) pendiente del suelo, (b) plantas nodrizas, (c) plantas sembradas y (d) plantación. Hábitats: (SA) suelo abierto, (BP) bajo dosel de pino y (VD) vegetación densa o del sotobosque.
Hábitat
De las 3600 semillas de pino, emergieron un total de 1254 plántulas, % de emergencia de plántulas =34.8%. Emergieron 464 plántulas a suelo abierto, 390 bajo el dosel de pino y 400 en vegetación densa. Sobrevivieron 14, 19 y 60 plántulas, a suelo abierto, bajo cobertura de pino y en vegetación densa, respectivamente, un total de 93 plántulas, % de supervivencia =7.2%. El análisis de varianza mostró diferencias significativas de supervivencia entre hábitats (F = 49.9; g. l. = 44, p < 0.01, figura 2-c). Las pruebas de comparación de promedio de Tukey y los intervalos de confianza muestran que las plántulas en vegetación densa presentaron el promedio más alto de supervivencia de plántulas (15.0a); a suelo abierto y bajo el dosel del pino se presentaron los promedios más bajos de supervivencia de plántulas: 3.0b y 4.8b, respectivamente. Los resultados indican que en la vegetación densa fue mayor la supervivencia de plantas, debido probablemente al efecto amortiguador del estrés ambiental de las plantas del sotobosque (tabla I).
Variación interanual en precipitaciones
Del total de 600 plántulas, sobrevivieron 85, % de supervivencia =14.2%. Mientras en 2010 las altas precipitaciones incrementaron la supervivencia de plantas (F= 179.1, g. l. = 1, p<0.01; figuras 1-b y 2-d), durante 2011 la tasa de supervivencia fue menor debido a la escasez de precipitaciones. En este ecosistema la humedad aprovechable del suelo puede cambiar entre años debido a la variación interanual en precipitaciones. Existieron diferencias estadísticas significativas en la supervivencia de plántulas (F= 67.4, g. l. =2; p<0.001). Las pruebas de comparación de promedios de Tukey y los IC gráficamente (figura 2-d) muestran mayores promedios de supervivencia de plantas en la vegetación densa VD-2010 (36.0a), seguido de BP-2010 (18.0b), SA-2010 (12.0b) y VD-2011 (11.0b). Los más bajos promedios de supervivencia de plantas se presentaron en los tratamientos BP-2011 (5.0c) y SA (3.0c). La interacción exposición a la lluvia x hábitats fue significativa (g.l = 2; p<0.001). Lo cual indica que las lluvias de 2010 (figura 1-b) y las plantas nodrizas del sotobosque (tabla I) incrementaron la supervivencia de plantas.
DISCUSIÓN
La mayor supervivencia de plántulas se registró en sitios con pendiente >30° (figura 2-a), debido probablemente a mayor conservación de humedad, menor exposición al sol y a temperaturas extremas. En suelos severamente degradados, el patrón reportado en plantaciones forestales de pino indica que a mayor pendiente mayor supervivencia, pero menor crecimiento (Gómez-Romero et al., 2012; Tejera y Davel, 2004). Las plantas nodrizas del sotobosque (tabla I) parecen incrementar la supervivencia del piñonero (figura 2-b) al amortiguar el estrés ambiental. La facilitación planta-planta se presenta desde la tundra a desiertos, en entornos alpinos hasta los bosques de lluvia (Soliveres et al., 2015; McIntire y Fajardo, 2014), y es común en ecosistemas áridos y semiáridos (Flores y Jurado, 2003) presentes en el noreste de México. La supervivencia de plantas sembradas fue mayor en vegetación densa (figura 2-c) lo cual coincide con los hallazgos de García y Jurado (2003; 2015) y García (2011). El efecto de la lluvia sobre la supervivencia de plántulas plantadas dependió del año, las lluvias torrenciales de 2010 (figura 1-b) en vegetación densa incrementaron la supervivencia de plántulas (figura 2-d). Un patrón de emergencia y supervivencia de plantas de matorral ha sido reportado en respuesta a las lluvias estacionales de verano-otoño en el noreste de México (García, 2011; García y Jurado, 2015). En áreas semiáridas con escasas lluvias, propensas a la sequía, como las del presente estudio, la supervivencia y establecimiento de plantas puede depender de eventos raros y extremos, como las intensas lluvias del verano-otoño de 2010.
CONCLUSIONES
(1) La pendiente > 30° incrementó la supervivencia de P. cembroides. (2) Las plantas nodrizas son un mecanismo de facilitación planta-planta debido a que reducen el estrés ambiental e incrementan la supervivencia de plántulas. (3) La supervivencia de plántulas emergidas en hábitats contrastantes fue mayor en vegetación densa y (4) las lluvias de 2010 incrementaron la supervivencia de plántulas en vegetación densa. Se sugieren programas de reforestación con plantas en sitios idóneos, aplicando hidrogel para la retención de humedad.
AGRADECIMIENTOS
A los estudiantes de la FA-La Chona, generaciones 2005-2014, por su ayuda en la colecta de datos, y a los directivos MC. Jesús Pedroza e Ing. Efrén Montaño, por las facilidades otorgadas. Promep/103.5/09/3905; y 103.5/10/6646.
* Universidad Autónoma de Nuevo León.
Contacto: jaimefgarcia@hotmail.com
REFERENCIAS
Conagua. (2009). Estadísticas. México: Comisión Nacional del Agua. p. 28.
Esparza, M. (2014). La sequía y la escasez de agua en México. Situación actual y perspectivas futuras. Secuencia. 195-219.
Flores, J., y Jurado, E. (2003). Are nurse-protégé interactions more common among plants from arid environments? Journal of Vegetation Science. 14: 911-916.
Frank, D., Reichstein, M., Bahn, M., et al. (2015). Effects of climate extremes on the terrestrial carbon cycle: concepts, processes and potential future impacts. Global Change Biology. 21(8): 2861-2880.
García, J.F., y Jurado, E. (2003). Influence of plant cover on germination in matorral in northeastern Mexico. Forest Ecology and Management. 177:11-16.
García-Pérez, J.F., Aguirre-Calderón, O., Estrada-Castillón, E., et al. (2007). Germinación y establecimiento de plantas nativas del matorral tamaulipeco y una especie introducida en un gradiente de elevación. Madera y Bosques. 13(1): 99-117.
García, J.F. (2011). Can environmental variation affect seedling survival of plants in northeastern Mexico? Archives of Biological Science Belgrade. 63: 731-737.
García, J.F., y Jurado, E. (2015). Is drought altering plant populations in the mountainous region of Northeastern Mexico? Acta Botanica Croatica. 74(1): 95-108.
Gobiet, A., Kotlarski, S., Beniston, M., et al. (2014). 21st century climate change in the European Alps-a review. Science of the Total Environment. 493: 1138-1151.
Gómez-Romero, M., Soto-Correa, J.C., Blanco-García, J.A., et al. (2012). Estudio de especies de pino para restauración de sitios degradados. Agrociencia. 46(8). 795-807.
Hughes, L. (2000). Biological consequences of global warming: Is the signal already apparent? Trends Ecol. Evol. 15(2): 56-61.
IPCC. (2013). Working Group I Contribution to the IPCC Fifth Assessment Report, Climate Change 2013: The Physical Science Basis. Cambridge University Press, New York, NY, USA.
Jurado, E., García, J.F., Flores, J., et al. (2006). Leguminous seedling establishment in Tamaulipan thorn scrub of Northeastern Mexico. Forest Ecol. Manage. 221: 133-139.
Lozano-Rivas, W.A. (2011). Determinación del número mínimo de observaciones en investigación, obviando las estimaciones de la varianza de datos. Revista de Didáctica Ambiental. 10: 54-61.
McIntire, E.J.B., y Fajardo, A. (2014). Facilitation as a ubiquitous driver of biodiversity. New Phytologist. 201: 403-416.
McLachlan, J.S., Clark, J.S., y Manos, P.S. (2005): Molecular Indicators of tree migración capacity under rapid climate change. Ecology. 86: 2088-2098.
Müller, L.L.B., Albach, D.C., y Zotz, G., (2017). “Are 3°C too much?”: thermal niche breadth in Bromeliaceae and global warming. Journal of Ecology. 105: 507-516.
Peterson, A.T., Egbert, S.L., Sánchez-Cordero, V., et al. (2002). Future projections for Mexican faunas under global climate change scenarios. Nature. 416: 626-629.
Soliveres, S., Maestre, F.T., Berdugo, M., et al. (2015). Missing link between facilitation and plant species coexistence: nurses benefit generally rare species more than common ones. J. Ecol. 103: 1183-1189.
Sokal, R.R., y Rohlf, F.J. (1995). Biometry: The Principles and Practice of Statistics in Biological Research. 3rd Edition, W.H. Freeman and Co., New York.
Tejera, L., y M. Davel. (2004). Establecimiento en pino Oregón en la Patagonia. Ficha técnica. Patagonia Forestal. 9(2): 9-12.
TABLA
Tabla I. Lista taxonómica y número de plántulas de vegetación del sotobosque encontradas en los veranos de 2009, 2011 y 2014. Las plántulas habitaban en un radio de 20 cm de P. cembroides a suelo abierto (SA) y bajo dosel del piñonero (BP) en vegetación densa (VD).