DINÁMICA DE NUTRIENTES EN SUELOS DEL MATORRAL ESPINOSO TAMAULIPECO
Perla Cecilia Rodríguez Balboa*, Marisela Pando Moreno*, Humberto González Rodríguez*, Israel Cantú Silva*, José Guadalupe Marmolejo Monsiváis*, Jonathan Lazcano Cortez*
CIENCIA UANL / AÑO 22, No.97 septiembre-octubre 2019
RESUMEN
La principal entrada de nutrientes al suelo se obtiene de la hojarasca que es de suma importancia en el entendimiento de los ciclos de nutrientes, la cual depende de tres factores: condiciones climáticas, calidad de la hojarasca y cantidad de microorganismos en el suelo. El papel de los microorganismos es importante en el funcionamiento de los ecosistemas, pues son los que se encargan de degradar la hojarasca y realizar la mineralización e inmovilización microbiana. El objetivo del presente documento es realizar una revisión de bibliografía fundamentalmente del ciclo de nutrientes en suelos del matorral espinoso tamaulipeco.
Palabras clave: ciclo de nutrientes en suelos, matorral espinoso tamaulipeco, condiciones climáticas, mineralización, hojarasca, microorganismos en el suelo.
ABSTRACT
Soil nutrients are mainly obtained from leaf litter, being of utmost importance the understanding of nutrient cycles. The nutrient cycle usually depends on three factors: climatic conditions, leaf litter quality and quantity of micro-organisms in the soil. The role of micro-organisms is of great importance in the functioning of ecosystems, as they usually degrade the leaf litter, and mineralize and immobilize the microbes. The objective of the present research is to make an extensive literature review on the cycle of soil nutrients in the thorny scrub of the Tamaulipas state.
Keywords: Soil nutrients cycle, thorny scrub tamaulipas, climate conditions, mineralization, leaf litter, soil micro-organisms.
El suelo es el mayor depósito de carbono, almacena 80% del carbono terrestre global, por lo que interviene en la captura del CO2 atmosférico (Yáñez-Díaz et al., 2017). El ecosistema denominado matorral espinoso tamaulipeco (MET) cuenta con una superficie de 200,000 km2 que abarca el noreste de México y sur de Texas (Alanís-Rodríguez et al., 2008). El suelo predominante en el MET es el denominado vertisol, el cual tiene una coloración gris oscuro, limoarcilloso con montmorillonita, tiene la capacidad de contraerse y expandirse en respuesta a diferentes cambios en el contenido de humedad del suelo (González-Rodríguez et al., 2011). La disponibilidad de nutrientes y la productividad de los ecosistemas dependen, en su mayoría, de la actividad microbiana presente en el suelo (Gama-Rodrigues, 2011), por tal motivo afecta las relaciones suelo-planta (García, Ramírez y Sánchez, 2012; Salete-Capellesso et al., 2016). La escasez de los nutrientes en el suelo afecta la productividad de los ecosistemas forestales (Cerón-Rincón y Aristizábal-Gutiérrez, 2012; Li, 2010) debido a que la descomposición de la materia orgánica a través de procesos físicos (lixiviación) y biológicos (actividad microbiana) determina el reciclado de nutrientes y su disponibilidad en el suelo para el aprovechamiento de las plantas (Fernández y Aragón, 2014; Fuentes-Molina y Rodríguez-Barrios, 2012; Pérez-Suárez, Arredondo-Moreno y Huber-Sannwald, 2012).
Los factores involucrados en la mineralización e inmovilización de nutrientes son la composición de la vegetación, las interacciones entre la materia orgánica, los microorganismos del suelo y las condiciones ambientales como precipitación y temperatura (Celentano et al., 2011; Gaspar-Santos et al., 2015).
FACTORES INVOLUCRADOS EN LA DINÁMICA DE NUTRIENTES
Los factores principales que controlan la dinámica de los nutrientes se describen a continuación:
Calidad de la hojarasca
La calidad de la hojarasca regula la capacidad del suelo para suministrar nutrientes, almacenar agua, liberar gases de efecto invernadero. Asimismo, las características fisicoquímicas del material orgánico, es decir, el contenido de agua, nitrógeno y demás contenido de nutrientes, en conjunto, influyen fuertemente en la liberación de nutrientes (Bradford et al., 2016; Hopkins y Dungait, 2010; Rawat, Nautiyal y Nautiyal, 2009).
Temperatura y precipitación
Las condiciones ambientales influyen en las características químicas de la hojarasca, regulando de esta manera el ciclo de nutrientes. En este sentido, los factores climáticos como la temperatura y precipitación influyen en la descomposición de la hojarasca (Marmolejo-Moncivais, Cantú-Ayala y Gutiérrez-Suárez, 2013), favoreciendo la presencia o ausencia de la actividad microbiana (Carnevele y Lewis, 2009; Wang, Wang y Huang, 2008).
El efecto combinado de precipitación y temperatura son considerados como factores importantes que condicionan la velocidad de descomposición, en periodos secos, cuando se retrasa la descomposición del material vegetal. Asimismo, las variables climáticas del suelo, como humedad y temperatura, están ampliamente relacionadas con la velocidad de degradación (Gallardo y Pino, 1988).
La temperatura óptima para que ocurra la descomposición de la materia orgánica oscila en un rango de 10-40°C. La actividad microbiana es nula en temperaturas de 0°C (Hopkins y Dungait, 2010). Ejemplo de lo anterior es la temperatura registrada en el MET, respectivamente para Linares, de 12.8°C mínima y 30.6°C máxima, lo que favorece la presencia de la actividad microbiana. En cambio, la precipitación registrada es de 629.9 mm (López-Hernández et al., 2013). De acuerdo con las variables ambientales presentes, en la región favorece la descomposición y la disponibilidad de nutrientes.
FACTORES DEL SUELO INVOLUCRADOS EN LA DINÁMICA DE NUTRIENTES
Los factores principales que controlan la dinámica de los nutrientes se describen a continuación:
Factores físicos
Textura
La textura juega un papel importante en el proceso de degradación de la hojarasca y mineralización de nutrientes. Los suelos arenosos tienen mayor mineralización debido a la aireación del suelo. Por otro lado, la arcilla tiene alta capacidad de retención de humedad, por lo que puede afectar procesos microbianos de mineralización e inmovilización. (Monsalve, Gutiérrez y Cardona, 2017).
La textura que se presenta en el matorral espinoso tamaulipeco es limoarcilloso, de acuerdo al triángulo de textura de la USDA con arena: 17.8%, limo: 41.2% y arcilla: 41.0% (Yáñez-Díaz et al., 2017).
Humedad del suelo
La humedad del suelo cumple una función importante en la descomposición de la hojarasca, al existir humedad los microorganismos se encuentran activos y realizan la descomposición y mineralización; sin embargo, en condiciones de desecación la mineralización es mínima e incluso provoca inmovilización de los nutrientes (Monsalve, Gutiérrez y Cardona, 2017).
Densidad aparente
La densidad aparente juega un papel importante en la mineralización de nutrientes, debido a que al presentarse suelos con mayor contenido de arcilla provoca que la retención de humedad sea más alta, disminuyendo el contenido de aire, lo que conlleva a la disminución de los microorganismos presentes en el suelo (Monsalve, Gutiérrez y Cardona, 2017). La densidad aparente en el matorral espinoso tamaulipeco es de 0.9 (g/cm3) (Yáñez-Díaz et al., 2017), este valor indica que la densidad aparente es baja y no representa un valor crítico en las propiedades físicas del suelo.
Factores químicos
pH
Es un indicador de la disponibilidad de los nutrientes, cuando el suelo tiene un pH cercano a la neutralidad o alcalino (pH≥6.5) provoca que los nutrientes sean no disponibles por las plantas debido a la abundancia de iones OH- provocando la precipitación de compuestos insolubles (Osorio, 2012). El valor de pH registrado en el MET es de 7.60 (Yáñez-Díaz et al., 2017), el valor indica que el pH es ligeramente alcalino. La disponibilidad de nutrientes es alta para el MET a excepción del Fe, Mn y P, los cuales presentan menor disponibilidad de acuerdo al valor del pH.
PAPEL DE LOS MICROORGANISMOS EN LA DEGRADACIÓN DE LA HOJARASCA
Diversidad microbiana
Los microorganismos en el suelo juegan un papel importante en la retención y liberación de nutrientes en los ecosistemas forestales, la relación entre microorganismos y plantas puede ser tanto de mutualismo como de competencia (Gallardo et al., 2009). Estos microrganismos se dividen en cinco categorías taxonómicas: algas, bacterias, hongos, protistas y virus. En el suelo están estrechamente asociados con las partículas, principalmente arcillas y materia orgánica, formando un microhábitat en el cual incluyen superficies interiores y exteriores de los agregados del suelo, por lo tanto, se puede afirmar que el suelo es heterogéneo con respecto a la distribución de los microoorganismos (Giri et al., 2005).
Estos microorganismos son de suma importancia debido a que juegan un papel en la descomposición y son considerados reservorios de nitrógeno, liberado con la muerte de los mismos (Celaya-Michel y Castellanos-Villegas, 2011). La diversidad de los microorganismos es más antigua que la existencia de las plantas y animales, por lo tanto, han tenido tiempo para evolucionar a diferentes formas (Giri et al., 2005).
Los hongos son capaces de descomponer la mayor parte de materia orgánica (Hopkins y Dungait, 2010), se presentan en el suelo en diferentes formas y tamaños, algunas especies forman colonias, mientras que otras son levaduras unicelulares. Las micorrizas son hongos que viven encima o en las raíces de las plantas, éstos incrementan la toma de agua y nutrientes (Crespo, 2013).
Las bacterias son organismos unicelulares y son los más numerosos en el suelo; uno de sus beneficios es que ayudan a las plantas a la toma de nutrientes. Los actinomicetos no son tan numerosos como las bacterias, tienen funciones importantes como descomponer la materia orgánica en humus y por tal motivo liberan nutrientes que son aprovechados por las plantas (Crespo, 2013).
Las algas son organismos que producen su propio alimento a través de la fotosíntesis, aparecen en el suelo después de las lluvias como una capa fina verde, la función principal es mejorar la estructura, aunque algunas especies, como las verdes azules, pueden fijar N que después liberan y puede ser absorbido por las plantas (Crespo, 2013). La actividad microbiana es pieza clave para el entendimiento de los procesos biológicos del suelo (Liu et al., 2000), por lo que cambios en el suelo pueden afectar a la productividad de las plantas (Crespo, 2013).
BALANCE ENTRE MINERALIZACIÓN E INMOVILIZACIÓN MICROBIANA
Mineralización de nitrógeno
La mineralización del nitrógeno es el proceso por el cual el nitrógeno orgánico del suelo es convertido por los microorganismos en compuestos inorgánicos (nitrato y amonio). El amonio puede ser fijado por la materia orgánica o las partículas de arcilla del suelo, se volatiliza como amoniaco y es absorbido por las plantas o los microorganismos. Estos compuestos inorgánicos son tomados por los microorganismos, incorporándolos a su biomasa, y se le conoce como inmovilización (Celaya-Michel y Castellanos-Villegas, 2011). El proceso de mineralización es similar para el resto de los nutrientes.
Los microorganismos del suelo son de gran importancia para la descomposición y mineralización del nitrógeno (Celaya-Michel y Castellanos-Villegas, 2011).
Métodos para evaluar la mineralización
Los métodos para calcular la mineralización representan un indicador de la cantidad de nitrógeno orgánico que cambia a inorgánico en un tiempo determinado, por lo cual se realizan incubaciones de algunas muestras de suelo, permitiendo calcular el amonio y nitrato que se mineraliza después de un tiempo determinado. Para obtener la tasa neta de mineralización in situ, se utiliza el método de las bolsas de polietileno cubiertas en el sitio a evaluar, se agrega suelo y se sellan, permanecen en el sitio por un periodo con las mismas condiciones de temperatura. Las bolsas pueden sufrir alteraciones causadas por los insectos o las raíces de las plantas, lo que contribuye a la pérdida de mineralización del nitrógeno. Sin embargo, si la bolsa no sufre daños durante el periodo de evaluación, no existirá lixiviación de nutrientes (Giri et al., 2005).
Inmovilización microbiana
La biomasa microbiana tiene la función de almacenamiento (inmovilización) y fuente (mineralización) de nutrientes, por lo que no sólo es utilizada como indicador de la calidad del suelo (Monsalve, Gutiérrez y Cardona, 2017). Se considera inmovilización a la absorción y asimilación microbiana de nutrientes que no son disponibles para las plantas (Taiz y Zeiger, 2002), al ocurrir la muerte de la biomasa microbiana se liberan los nutrientes que se encontraban inmovilizados.
IMPORTANCIA DE LA HOJARASCA EN LA DINÁMICA DE LOS NUTRIENTES
Los primeros estudios sobre la hojarasca se realizaron en la década de 1960, debido al papel que juega en la comprensión de los nutrientes. La hojarasca es la principal fuente de nutrientes y materia orgánica en la capa del humus del suelo en el matorral espinoso tamaulipeco (González-Rodríguez et al., 2017). La hojarasca constituye la principal entrada de los nutrientes al suelo y es por ello que forma parte del punto clave del reciclado de nutrientes y materia orgánica (Gutiérrez-Vázquez et al., 2012; Crespo, 2015). Los nutrientes más comunes en la hojarasca son N, P, K y Ca, y en la mayoría de los casos el N es el nutriente más abundante (Gutiérrez-Vázquez et al., 2012).
La hojarasca está compuesta por hojas, ramas, inflorescencias, frutos, entre otros, los cuales son depositados al suelo (Gutiérrez-Vázquez et al., 2012; Crespo, 2015). La hojarasca, al caer al suelo, tiene una atribución importante en su formación (Triadiati et al., 2011), gracias a la circulación de nutrientes y materia orgánica que ocurre con la descomposición (Kumar y Tewari, 2014).
La descomposición de la hojarasca se divide en cuatro etapas: trituración, el fraccionamiento de los tejidos; lixiviación, la pérdida de los compuestos más solubles mediante corrientes de agua; catabolismo (se incluye la mineralización y la humificación), la transformación que realizan los microorganismos de los compuestos orgánicos, es decir, transforman los nutrientes para que estén disponibles para las plantas y para la humificación, es la formación de la materia orgánica por los microorganismos (Álvarez-Sánchez, 2001). El ciclo de nutrientes y la descomposición de la hojarasca son procesos complejos que se dividen en tres pasos: descomposición, mineralización y humificación (Mishra y Kumar, 2016; Sánchez 2008).
IMPORTANCIA DE LA MATERIA ORGÁNICA EN LA DINÁMICA DE LOS NUTRIENTES
La materia orgánica está conformada por componentes ricos en nitrógeno, carbono, fósforo y agua, principalmente, los cuales propician energía y nutrimentos necesarios para el desarrollo y metabolismo de los microorganismos encargados de la descomposición (Ferrera-Cerato y Alarcón, 2001).
En la materia orgánica se encuentra la mayor cantidad de nitrógeno, pero no está disponible para las plantas y mediante el proceso de descomposición y mineralización el nitrógeno es transformado a compuestos inorgánicos que lo regresan disponible (Celaya-Michel y Castellanos-Villegas, 2011). La descomposición de la materia orgánica es un proceso que permite reciclar nutrientes (Shi, 2011) que no están disponibles para las plantas, esta tarea es llevada a cabo en su totalidad por los microorganismos que se encuentran en el suelo (Celaya-Michel y Castellanos-Villegas, 2011). Durante el proceso de descomposición primero se libera la fracción lábil (azúcares y proteínas) y después la fracción recalcitrante, la cual es de lenta descomposición (ligninas y fenoles) (Sánchez et al., 2008).
La calidad de la materia orgánica es importante debido al papel que desempeña en los procesos funcionales que se llevan a cabo en los ecosistemas forestales (López-Hernández et al., 2013). La entrada de la materia orgánica al suelo está determinada por la caída de hojarasca, tanto producción como descomposición debido a que son piezas clave en el reciclado de nutrientes (Cantú-Silva 2013).
Algunas funciones que tiene la materia orgánica en el suelo se mencionan a continuación (García-Leaños, 2008):
- Es la fuente de crecimiento de las bacterias, las cuales liberan sustancias que ayudan en la formación de agregados para hacer más estable el suelo.
- Retiene minerales, lo que reduce su arrastre y pérdida.
- Reduce la erosión causada por la lluvia.
- Incrementa la penetración de las raíces.
- Regula la temperatura y humedad del suelo.
IMPORTANCIA DE LOS NUTRIENTES EN EL SUELO
Los nutrientes son elementos que se derivan del suelo y son esenciales para las plantas, de acuerdo a su concentración y a los requerimientos necesarios para el crecimiento de las mismas. Las plantas requieren 17 elementos esenciales, los cuales se dividen en macronutrientes y micronutrientes. Cada nutriente tiene funciones específicas en las plantas (Cogger y Brown, 2016; Kyrkby y Römheld, 2007). La absorción de nutrientes por las plantas está inicialmente limitada por las tasas de descomposición y de mineralización, la solubilidad mineral, la capacidad de intercambio catiónico y la competitividad de los microorganismos (Bosco-Imbert, Blanco y Castillo, 2004). Se estima que durante la descomposición de la hojarasca la cantidad de nutrientes liberados equivalen a 70-90% del total de los requeridos por las plantas (Sánchez et al., 2008).
Macronutrientes
Los macronutrientes son requeridos en grandes cantidades por las plantas, de ahí su nombre. Los nueve principales nutrientes son: nitrógeno, carbono, fósforo, potasio, calcio, magnesio, azufre (Cogger y Brown, 2016; Kyrkby y Römheld, 2007), hidrógeno y oxígeno (Rodríguez y Flórez, 2004).
Micronutrientes
Los micronutrientes son requeridos en cantidades menores; asimismo, son considerados elementos esenciales para el crecimiento y desarrollo de las plantas. Los micronutrientes son: hierro, manganeso, boro, zinc, cobre, molibdeno, cloro (Cogger y Brown, 2016; Kyrkby y Römheld, 2007), níquel (Rodríguez y Flórez, 2004).
PRINCIPALES CICLOS DE NUTRIENTES
Ciclo del nitrógeno
El nitrógeno es un elemento esencial requerido para la degradación de la materia orgánica y el crecimiento microbiano. Si la materia orgánica tiene alto contenido de nitrógeno, los microorganismos tienen suficiente sustrato para inducir mayor mineralización, satisfaciendo totalmente sus necesidades de N, es por ello que no representa un factor limitante. Por otro lado, si el contenido de N es bajo, la descomposición de la materia orgánica disminuye drásticamente y la mineralización de carbono orgánico dependerá de las fuentes nitrogenadas (Ferrera-Cerato y Alarcón, 2001).
El ciclo del nitrógeno es de suma importancia pues mediante éste los nutrientes son liberados y adquiridos por la materia orgánica (Cogger y Brown, 2016), se incluyen cuatro procesos: fijación del nitrógeno, mineralización, nitrificación y desnitrificación (Mao, Yannarell y Mackie, 2011). Este elemento puede ser absorbido por las raíces en tres formas: como N orgánico, amonio o nitrato. La mineralización y nitrificación en el suelo son procesos lentos, por lo que suelen predominar las formas orgánicas y reducidas, disminuyendo la abundancia en el suelo en el sentido: N orgánico soluble > amonio > nitrato (Escudero y Mediavilla, 2003).
Ciclo del carbono
La descomposición de la hojarasca es importante en el ciclo del carbono (Zhang et al., 2008; De Paz et al., 2017). El carbono desempeña un papel central en el control del reciclaje del nitrógeno (Cantú-Silva y Yáñez-Díaz, 2018).
Para conocer la velocidad con que la materia orgánica se mineraliza, se utiliza la relación entre carbono y nitrógeno, la cual indica la tasa de nitrógeno disponible para las plantas, por tal motivo, al obtener valores de 10 a 14 corresponden a una mineralización rápida, lo que favorece a que los microorganismos y las plantas cuenten con los nutrientes suficientes (Gamarra-Lezcano et al., 2017).
Por tal motivo los valores altos de la relación carbono y nitrógeno indican que la hojarasca se descompone más lento, a diferencia de valores bajos en los que la degradación es más rápida, debido principalmente la calidad de la hojarasca (Álvarez-Sánchez, 2001).
El C orgánico del suelo resulta del balance entre la incorporación del material orgánico al suelo y la salida del C del suelo en forma de CO2 a la atmósfera, es emitido desde el suelo a la atmósfera, pero no sólo es producido por la mineralización de la materia orgánica, sino que es generado también por el metabolismo de las raíces de las plantas (Martínez, Fuentes y Acevedo, 2008).
Ciclo del azufre
El azufre es un elemento necesario para las plantas en menor proporción que el N, pero es tan importante como el ya mencionado, el proceso de asimilación ocurre al pasar de las formas oxidadas a reducidas del azufre, este cambio de oxidación del azufre es una actividad realizada en su mayoría por los microorganismos del suelo. El azufre se encuentra en la atmósfera como dióxido de azufre (SO2), por lo que no se encuentra disponible para las plantas, es por ello que en el ciclo del azufre se convierte en sulfato de azufre (SO4-2), que es absorbido por las raíces de las plantas e incorporado al aminoácido cisteína (Benavides, 1998).
Ciclo del fósforo
El fósforo es un elemento que proviene de las apatitas (fosfato de calcio natural más abundante en la superficie terrestre, es por ello que son consideradas la principal fuente de fósforo) (Fernigrini, Cámara y Oliva, 2008). El fosfato es liberado mediante los procesos de lixiviación, meteorización y erosión. Es absorbido por las plantas y los microorganismos del suelo, por lo que es incorporado en la materia orgánica y sedimentos, para depositarse nuevamente en forma de mineral no disponible para la asimilación. El fósforo inorgánico (H2PO4- y HPO42-) por lo general es fijado fuertemente en forma de fosfatos de Mg2+, Ca2+, Al3+ y Fe2+. La mineralización de los compuestos orgánicos (ácidos nucleicos, fosfolípidos y ésteres) del fósforo permite el reciclaje del elemento para que regrese al suelo y permita formar parte del ciclo. A diferencia del ciclo del nitrógeno, el ciclo del fósforo no incluye cambios en los compuestos inorgánicos, es tomado del suelo por las plantas y retorna a través de los residuos vegetales y animales (Cerón-Rincón y Aristizábal-Gutiérrez, 2012).
Ciclo del potasio
El potasio es un elemento esencial, por lo que las plantas necesitan cantidades elevadas similares al nitrógeno, se obtiene del suelo y proviene de la meteorización de minerales, mineralización de los residuos orgánicos y en algunos casos de los abonos o fertilizantes. Los compuestos de arcilla son la principal fuente de potasio en el suelo. Se encuentra disponible en la solución del suelo, por lo que es absorbido por las plantas de manera inmediata. El ciclo del potasio en el suelo consiste en la adsorción y desorción de potasio intercambiable de la solución del suelo, el cual es fijado o liberado en las arcillas y se cristaliza o meteoriza en potasio estructural (Conti, 2002).
El potasio de la materia orgánica pasa a la solución del suelo, asimismo, se pueden aplicar fertilizantes de potasio. Las pérdidas de potasio en la solución del suelo se presentan con la erosión del suelo y el potasio lixiviado (Conti, 2002).
FUNCIONES DE LOS NUTRIENTES
Las funciones de los macro y micronutrientes en las plantas se ilustran en la siguiente tabla:

Tabla I. Funciones principales de macro y micronutrientes y la forma disponible de asimilación por las plantas entre paréntesis (Taiz y Zeiger, 2002; Salisbury y Ross, 1985).
ISÓTOPOS EN EL CICLO DE NUTRIENTES
Los isótopos estables son los elementos con el mismo número atómico, pero diferente peso atómico. Se utilizan para estudiar los efectos del uso del suelo sobre las características de los ecosistemas (Boutton, Archer y Midwood, 1999). Se tienen estudios con isótopos estables de H, C, N, O y S en el sur de Texas y de ahí su importancia en los ecosistemas para registrar cambios de vegetación de pastos a bosques.
DISCUSIÓN
Con base en la bibliografía consultada sobre la importancia del ciclo de nutrientes y el papel que juega la hojarasca como la principal entrada de nutrientes en el suelo, resulta evidente la importancia de los microrganismos, los cuales cumplen una función importante para degradar la hojarasca y liberar nutrientes que son aprovechados por las plantas. Existen diversos estudios relacionados al aporte de nutrientes en el ecosistema del matorral espinoso tamaulipeco.
El suelo forma parte del reservorio de carbono, el cual funciona como sumidero de CO2, en este sentido, Cantú y Yáñez (2018) evaluaron el contenido de carbono orgánico del suelo, nitrógeno total y se estimó la relación de C/N en dos profundidades de 0-5 y 5-30 cm, siendo la primera en la que se presentó mayor cantidad de los contenidos de carbono orgánico, nitrógeno total y relación de C/N (Cantú-Silva y Yáñez-Díaz, 2018). Asimismo, Yáñez et al. (2017) estudiaron las variaciones diurnas y estacionales en el flujo del CO2 en vertisoles bajo cuatro sistemas de uso de suelo, obteniendo que en el sistema de matorral se registró el valor más alto de respiración con 6.17 µmol CO2 m-2 s-1 por la mañana y 8.4 µmol CO2 m-2 s-1 por la tarde, que indica la emisión de CO2, lo que podría afectar el balance del carbono (Yáñez-Díaz et al., 2017). El contenido de carbono en especies arbóreas y arbustivas en diferentes componentes de la biomasa fue evaluado por Yerena et al. (2012), quienes encontraron valores de 48.14% en hojas, 45.93% en tallos, 45.73% en ramas, 45.70% en ramillas y 42.08% en corteza (Yerena-Yamallel et al., 2012).
Con la finalidad de conocer el aporte de nutrientes que se obtiene en la producción de la hojarasca, López et al. (2013) analizaron durante un año la producción de hojarasca y el aporte de macro y micronutrientes, los resultados indicaron que en invierno se depositó la mayor cantidad de producción foliar y, asimismo, el aporte de nutrientes (López-Hernández et al., 2013). Del mismo modo, López et al. (2015) evaluaron la producción de hojarasca y depósito potencial de nutrientes en el componente hojas, el depósito de nutrientes fue más alto en otoño e invierno (López-Hernández et al., 2015).
Mientras que Cantú et al. (2013) determinaron el retorno potencial de nutrientes por los componentes secundarios de la hojarasca; las estructuras reproductivas son el componente de mayor importancia en el depósito de nutrientes, además encontraron diferencias entre los sitios de estudio, las cuales se deben principalmente a la composición de la vegetación, la calidad de hojarasca, las condiciones edáficas y las variables climáticas (Cantú-Silva 2013).
ESTUDIOS DE NUTRIENTES EN EL MATORRAL ESPINOSO TAMAULIPECO (MET)
A continuación, se describen algunos estudios relacionados al aporte de nutrientes en el MET:
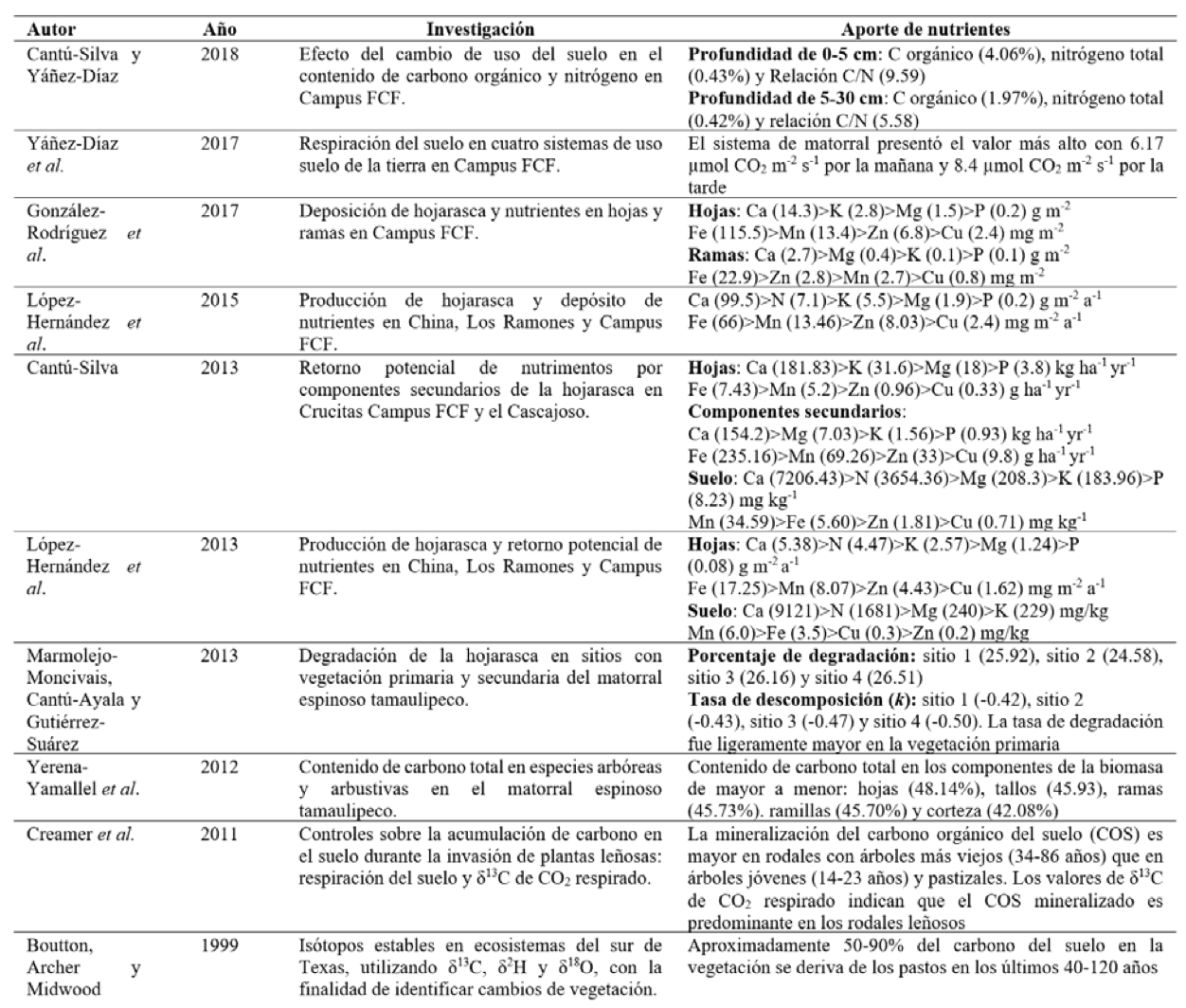
Tabla II. Estudios relacionados al aporte de nutrientes en el MET.
Por otro lado, la importancia de la tasa de descomposición es de suma importancia y es por ello que Marmolejo et al. (2013) realizaron un estudio para identificar la tasa de descomposición de la hojarasca en cuatro sitios con vegetación primaria y secundaria, los resultados indicaron que la tasa de degradación fue mayor en la vegetación primaria (Marmolejo-Moncivais, Cantú-Ayala y Gutiérrez-Suárez, 2013).
Los microorganismos son importantes en el proceso de mineralización de nutrientes, ante ello Celaya y Castellanos (2011) evaluaron la tasa de mineralización del nitrógeno, un indicador de la disponibilidad del nitrógeno en zonas áridas y semiáridas, en el cual concluyeron que, debido a la poca vegetación, la mineralización se presenta de manera heterogénea causada por las condiciones del microclima y el aporte de materia orgánica, por tal razón los valores de mineralización son mayores bajo el dosel de árboles y arbustos, principalmente en leguminosas (Celaya-Michel y Castellanos-Villegas, 2011). Es viable conocer la tasa de mineralización de los nutrientes, por ejemplo, el nitrógeno, empleando la metodología recomendada por Celaya y Castellanos.
Diversos estudios señalan que las condiciones ambientales influyen en la descomposición de la hojarasca debido a que inhiben o favorecen la presencia de microorganismos en el suelo, Bradford et al. (2016) mencionan que desde hace tiempo se ha considerado el clima como el principal factor que controla la descomposición, pero indican que los rasgos propios de la hojarasca pueden predominar en la descomposición, por lo que proponen realizar estudios más precisos para identificar cuáles son los principales factores que la controlan (Bradford et al., 2016).
Con respecto al uso de isótopos, Boutton et al. (1999) evaluaron ecosistemas del sur de Texas, utilizando isótopos estables de δ13C, δ2H y δ18O, con la finalidad de identificar cambios de vegetación, y encontraron que durante los últimos 40-120 años se presentó un cambio de vegetación de pastos a bosques, por lo que afirman que cerca de 50-90% del carbono del suelo se deriva de los pastos (Boutton, Archer y Midwood, 1999). Asimismo, Creamer et al. (2011) cuantificaron controles sobre la acumulación de carbono en el suelo durante la invasión de plantas leñosas: respiración del suelo y δ13C de CO2 respirado, en el cual utilizaron fracción del suelo para investigar la degradación microbiana de la materia orgánica, para identificar por qué la mineralización del carbono orgánico (COS) es mayor en rodales con árboles más viejos. Los valores de δ13C de CO2 respirado indican que el COS mineralizado es predominante en los rodales leñosos debido a la fracción de luz (Creamer et al., 2011).
CONCLUSIONES
La disponibilidad de nutrientes afecta las relaciones entre suelo-planta, depende principalmente de las condiciones climáticas, la composición de la vegetación y los microorganismos del suelo.
Con la finalidad de conocer el aporte de nutrientes al suelo mediante la cuantificación de la producción de la hojarasca y la tasa de velocidad de degradación se ha logrado entender la dinámica de los nutrientes.
Se recomienda estudiar ampliamente los organismos que habitan en el suelo, debido a que los estudios son limitados, de igual manera son piezas clave en la descomposición de la hojarasca por la importancia que tienen de mineralizar e inmovilizar los nutrientes.
Existen diversos estudios relacionados con la acumulación y descomposición de la hojarasca en diferentes ecosistemas, pero desafortunadamente en el matorral espinoso tamaulipeco existen pocos trabajos que relacionen a los microorganismos y las variables climáticas en el proceso de descomposición y liberación de nutrientes.
Por lo tanto, se sugiere realizar investigaciones en las cuales se determinen las tasas de descomposición, mineralización, disponibilidad de nutrientes, enfocados a las características físicas y químicas del suelo, variables ambientales y microorganismos en el suelo, por lo que serán de gran utilidad en el matorral espinoso tamaulipeco.
El conocimiento integral de todos estos factores involucrados en la descomposición de la hojarasca permitirá entender la dinámica de los nutrientes, uno de los procesos más importantes en el ecosistema. Es de suma importancia conocer lo que sucede en los ecosistemas para poder tomar decisiones acertadamente y no poner en riesgo los recursos naturales.
* Universidad Autónoma de Nuevo León.
Contacto: perla.rodriguezbl@uanl.edu.mx
REFERENCIAS
Alanís-Rodríguez, E., Jiménez-Pérez, J., Aguirre-Calderón, O., et al. (2008). Efecto del uso del suelo en la fitodiversidad del matorral espinoso tamaulipeco. Revista Ciencia UANL. 11(1): 56-62.
Álvarez-Sánchez, J. (2001). Descomposición y ciclo de nutrientes en ecosistemas terrestres de México. Acta Zoológica Mexicana. 1: 11-nos27.
Benavides, A. (1998). El azufre de las plantas. Departamento de Horticultura. UAAAN.
Bosco-Imbert, J., Blanco, J.A., y Castillo, J.F. (2004). Gestión forestal y ciclos de nutrientes en el marco del cambio global. En: Valladares, F. Ecología del bosque mediterráneo en un mundo cambiante. 479-506.
Boutton, T.W., Archer, S.R., y Midwood, A.J. (1999). Stable isotopes in ecosystem science: structure, function and dynamics of subtropical savanna. Rapid Communications in Mass Spectrometry. 13: 1263-1277.
Bradford, M.A., Berg, B., Maynard, D.S., et al. (2016). Understanding the dominant controls on litter decomposition. Journal of Ecology. 104: 229-238.
Cantú-Silva, I., Sánchez-Castillo, L.R.M., González-Rodríguez, H., et al. (2013). Retorno potencial de nutrimentos por componentes secundarios de la hojarasca en comunidades vegetales de Nuevo León. Revista Mexicana de Ciencias Forestales. 4(17): 138-155.
Cantú-Silva, I., y Yáñez-Díaz, M.I. (2018). Efecto del cambio de uso de suelo en el contenido del carbono orgánico y nitrógeno del suelo. Revista Mexicana de Ciencias Forestales. 9(45): 122-151.
Carnevele, N.J., y Lewis, J.P. (2009). Leaf litter quality and litter decomposition of woody species in a seasonal subtropical forest (Argentina). Ecotrópica. 15: 23-33.
Celentano, D., Zahawi, R.A., Finegan, B., et al. (2011). Restauración ecológica de bosques tropicales en Costa Rica: efecto de varios modelos en la producción, acumulación y descomposición de hojarasca. Revista de Biología Tropical. 59(3): 1323-1336.
Celaya-Michel, H., y Castellanos-Villegas, A.E. (2011). Mineralización de nitrógeno en el suelo de zonas áridas y semiáridas. Terra Latinoamericana. 29(3): 343-356.
Cerón-Rincón, L.E., y Aristizábal-Gutiérrez, F.A. (2012). Dinámica del ciclo del nitrógeno y fósforo en suelos. Revista Colombiana de Biotecnología. 14(1): 285-295.
Cogger, C., y Brown, S. (2016). Soil formation and nutrient cycling. Sowing Seeds in the City. Springer Netherlands. 25-52. DOI 10.1007/978-94-017-7453-6_2.
Conti, M.E. (2002). Dinámica de la liberación y fijación de potasio en el suelo. Universidad de Buenos Aires, 14 pp.
Creamer, C.A., Filley, T.R., Boutton, T.W., et al. (2011). Control son soil carbon accumulation during woody plant encroachment: Evidence from physical fractionation, soil respiration, and δ13C of respired CO2. Soil Biology & Biochemistry. 43: 1678-1687.
Crespo, G. (2013). Funciones de los organismos del suelo en el ecosistema de pastizal. Revista Cubana de Ciencia Agrícola. 47(4): 329-334.
Crespo, G. (2015). Factores que influyen en el reciclaje de nutrientes en pastizales permanentes, avances en el desarrollo de su modelación. Revista Cubana de Ciencia Agrícola. 49(1): 1-10.
De Paz, M., Gobbi, M.E., Raffaele, E., et al. (2017). Litter decomposition of woody species in shrublands of NW Patagonia: how much do functional groups and microsite conditions influence decomposition? Plant Ecology. 218: 699-710.
Escudero, A., y Mediavilla, S. (2003). Dinámica interna de los nutrientes. Ecosistemas. 12(1): 1-8.
Fernández, R.D., y Aragón, R. (2014). Descomposición de hojarasca de las especies leñosas nativas y exóticas más abundantes del pedemonte de las Yungas, Tucumán, Argentina. Ecología Austral. 24: 286-293.
Fernigrini, P., Cámara, O.R., y Oliva, F.Y. (2008). Síntesis y caracterización de distintas apatitas. Su utilización como biomaterial. 2do Encuentro de Jóvenes Investigadores en Ciencia y Tecnología de Materiales. 5 pp.
Ferrera-Cerato, R., y Alarcón, A. (2001). La microbiología del suelo en la agricultura sostenible. Ciencia Ergo Sum. 8(2): 175-183.
Fuentes-Molina, N., y Rodríguez-Barrios, J.A. (2012). Eficiencia en el retorno potencial de nutrientes vía hojarasca de un bosque tropical de ribera. Sierra Nevada de Santa Marta-Colombia. Acta Biológica Colombiana. 17(1): 51-66.
Gallardo, A., Covelo, F., Morillas, L. et al. (2009). Ciclos de nutrientes y procesos edáficos en los ecosistemas terrestres: especificidades del caso mediterráneo y sus implicaciones para las relaciones suelo-planta. Ecosistemas. 18(2): 4-19.
Gallardo, A., y Pino, J. (1988). Importancia del medio físico en la descomposición de la hoja de especies arbóreas. Lagascalia. 15: 541-547.
Gama-Rodrigues, A.C. (2011). Soil organic matter, nutrient cycling and biological dinitrogen-fixation in agroforestry systems. Agroforestry Systems. 81: 191-193.
Gamarra-Lezcano, C.C., Díaz-Lezcano, M.I., Vera-de Ortiz, M., et al. (2017). Relación carbón-nitrógeno en suelos de sistemas silvopastoriles del Chaco paraguayo. Revista Mexicana de Ciencias Forestales. 9(46): 4-26.
García, Y., Ramírez, W., y Sánchez, S. (2012). Indicadores de la calidad de los suelos: una nueva manera de evaluar este recurso. Pastos y Forrajes. 35(2): 125-138.
García-Leaños, M.L. (2008). Materia orgánica: la sangre de la Tierra. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP). 1-6.
Gaspar-Santos, E.S., González-Espinosa, M., Ramírez-Marcial, N., et al. (2015). Acumulación y descomposición de hojarasca en bosques secundarios del sur de la Sierra Madre de Chiapas, México. Bosque. 36(3): 467-480.
Giri, B., Giang, P.H., Kumari, R., et al. (2005). Microbial Diversity in Soils. In: Buscot, F. and Varma, A. (eds.) Soil Biology. Microorganisms in Soils: Roles in Genesis and Functions. 3: 19-55.
González-Rodríguez, H., Domínguez-Gómez, T.G., Cantú-Silva, I., et al. (2011). Literfall deposition and leaf litter nutrient return in different locations at Northeastern Mexico. Plant Ecology. 212: 1747-1757.
González-Rodríguez, H., Ramírez-Lozano, R.G., Cantú-Silva, I., et al. (2017). Deposition of litter and nutrients in leaves and twigs in different plant communities of northeastern Mexico. Journal of Forest Research. Disponible en: https://doi.org/10.1007/s11676-017-0553-x.
Gutiérrez-Vázquez, M.H., Méndez-González, J., Flores-López, C., et al. (2012). Caída de hojarasca en plantaciones de Pinus greggii Engelm. y Pinus cembroides Zucc., en Coahuila, México. Revista Fitotecnia Mexicana. 35(2): 123-133.
Hopkins, D., y Dungait, J.A.J. (2010). In: Dixon, G.R., y Tilston, E.L. (eds.) Soil Microbiology and Sustainable Crop Production, Springer Science + Business Media B.V., 59-80.
Kumar, S., y Tewari, L.M. (2014). Pattern of litter fall and litter decomposition in a Quercus leucotrichophora A. Camus forest in Kumaun Himalaya. International Journal of Biodiversity and Conservation. 6(1): 108-114.
Kyrkby, E., y Römheld, V. (2007). Micronutrients in plants physiology: functions, uptake and mobility. Proceedings 543. The International Fertilizer Society.
Li, X., Hu, Y., Han, S., et al. (2010). Litterfall and litter chemistry change over time in an old-growth temperate forest, northeastern China. Annals of Forest Science. 67: 206-213
Liu, X., Lindemann, W., Whitford, W., et al. (2000). Microbial diversity and activity of disturbed soil in the northern Chihuahuan Desert. Biology and Fertility of Soils. 32: 243-249.
López-Hernández, J.M., González-Rodríguez, H., Ramírez-Lozano, R.G., et al. (2013). Producción de hojarasca y retorno potencial de nutrientes en tres sitios del estado de Nuevo León, México. Polibotánica. 35: 41-64.
López-Hernández, J.M., González-Rodríguez, H., Ramírez-Lozano, R.G., et al. (2015). Producción de hojarasca y depósito potencial de nutrientes de las hojas en el matorral espinoso tamaulipeco. Revista Mexicana de Ciencias Forestales. 6(30): 74-89.
Mao, Y., Yannarell, A., y Mackie, R. (2011). Changes in N-transforming archaea and bacteria in soil during the establishment of bioenergy crops. PLoS ONE. 6(9): 1-12.
Marmolejo-Moncivais, J.G., Cantú-Ayala, C.M., y Gutiérrez-Suárez, M.A. (2013). Degradación de la hojarasca en sitios con vegetación primaria y secundaria del matorral espinoso tamaulipeco. Revista Mexicana de Ciencias Forestales. 4(17): 174-181.
Martínez, H.E., Fuentes, E.J.P., y Acevedo, H.E. (2008). Carbono orgánico y propiedades del suelo. Revista de la Ciencia del Suelo y Nutrición Vegetal. 8(1): 68-96.
Mishra, G., y Kumar, R. (2016). Plant litter decomposition: drivers insight to the ecological process. European Journal of Biological Research. 6(3): 176-185.
Monsalve, C.O.I., Gutiérrez, D.J.S., y Cardona, W.A. (2017). Factores que intervienen en el proceso de mineralización de nitrógeno cuando son aplicadas enmiendas orgánicas al suelo. Una revisión. Revista Colombiana de Ciencias Hortícolas. 11(1): 200-209.
Osorio, N.W. (2012). pH del suelo y disponibilidad de nutrientes. Manejo Integral del Suelo y Nutrición Vegetal. 1(4): 1-4.
Pérez-Suárez, M., Arredondo-Moreno, J.T., y Huber-Sannwald, E. (2012). Early stage of single and mixed leaf-litter decomposition in semiarid forest pine-oak: the role of rainfall and microsite. Biogeochemistry. 108: 245-258.
Rawat, N., Nautiyal, B.P., y Nautiyal, M.C. (2009). Litter production pattern and nutrients discharge from decomposing litter in an Himalayan alpine ecosystem. New York Science Journal. 2(6): 54-67.
Rodríguez, S.M., y Flórez, R.V.J. (2004). Elementos esenciales y beneficiosos. Tecnologías y Programación en Agroplasticultura. 25-36.
Salete-Capellesso, E., Luana-Scrovonski, K., Zanin, E.M., et al. (2016). Effects of forest structure on litter production, soil chemical composition and litter-soil interactions. Acta Botanica Brasilica. 1-7.
Salisbury, F.B., y Ross, C.W. (1985). Plant Physiology. Belmont (CA):Wadsworth Publishing Company, 96-113.
Sánchez, S., Crespo, G., Hernández, M., et al. (2008). Factores bióticos y abióticos que influyen en la descomposición de la hojarasca en pastizales. Pastos y Forrajes. 31(2): 99-118.
Shi, W. (2011). Agricultural and Ecological significance of soil enzymes: soil carbon sequestration and nutrient cycling. In: Shukla, G. and Varma, A. (eds.). Soil Enzymology, Siol Biology 22, Springer-Verlag Berlin Heidelberg. 43-60.
Swift, M.J., y Anderson, J.M. (1989). Decomposition. En: Lieth, H., Werger, M.J.A. (Eds.): Tropical Rain Forest Ecosystems. Biogeographical and Ecological Studies Elsevier. New York, U.S.A. 547-567.
Taiz, L., y Zeiger, E. (2002). Plant physiology. The Benjamin/Cummings Publishing Company. California. 67-86.
Triadiati, S., Tjitrosemito, E., Guhardja, E., et al. (2011). Literfall production and leaf-litter decomposition at natural forest and cacao agroforestry in Central Sulawesi, Indonesia. Asian Journal of Biological Sciences. 4(3): 221-234.
Yáñez-Díaz, M.I., Cantú-Silva, I., González-Rodríguez, H., et al. (2017). Respiración del suelo en cuatro sistemas de uso de la tierra. Revista Mexicana de Ciencias Forestales. 8(42): 123-149.
Yerena-Yamallel, J.I., Jiménez-Pérez, J., Aguirre-Calderón, O.A., et al. (2012). Contenido de carbono total en los componentes de especies arbóreas y arbustivas en áreas con diferente uso, en el matorral espinoso tamaulipeco, en México. Bosque. 33(2): 145-152.
Wang, Q., Wang, S., y Huang, Y. (2008). Comparisons of litterfall, litter decomposition and nutrient return in a monoculture Cunninghamia lanceolata and a mixed stand in southern China. Forest Ecology and Management. 255: 1210-1218.
Zhang, D., Hui, D., Lou, Y., et al. (2008). Rates of litter decomposition in terrestrial ecosystems: global patterns and controlling factors. Journal of Plant Ecology. 1(2): 85-93.
Recibido: 01/03/2019
Aceptado: 07/06/2019