Biomineralización en una concha del Cretácico: Exogyra ponderosa
Milton Gómez-Ruiz*, Karen A. Zuñiga-Doria*, Elizabeth Chacón Baca*
CIENCIA UANL / AÑO 20, No. 85 julio-septiembre 2017
El nacimiento de Venus (Nascita di Venere), encargado por algún miembro no identificado de la familia de los Médicis y plasmado por Botticelli entre 1484 y 1485, es una de las pinturas renacentistas que han inmortalizado no sólo un derroche de hermosura y gracia femeninas, sino también la belleza de las conchas secretadas por algunos moluscos bivalvos, siempre presentes en la ciencia, en la literatura, en el arte y en la mitología (figura 1).

Figura 1. El nacimiento de Venus, de Sandro Botticelli, está plasmado en un gran lienzo que se encuentra actualmente en la colección de la Galería de Uffizi de Florencia, Italia (Uffizi, 2017).
Se dice que entre los aspectos notables de este lienzo de gran tamaño, destaca la posición antinatural y casi volátil de Venus, la diosa del amor, emergiendo a partir de una concha nacarada (probablemente un callo de hacha), en compañía de otras deidades como Céfiro (dios del viento) su consorte Aura, la ninfa de la Brisa y la Primavera quien, del otro lado, cubre a Venus con una capa floreada (Vasari, 1986). El abrazo entre Céfiro y Aura estaría representando la unión de los mundos espiritual y físico. Aunque esta obra ha sido objeto de diversas interpretaciones, el hecho de que Venus cubra su desnudez con la mano que posa sobre su exuberante cabellera rubia es considerada por algunos como un elemento diagnóstico de la filosofía neoplatónica (Acidini, 2009), en la cual dicha desnudez se representa como el ideal de la humanidad, y donde la belleza espiritual se conforma a partir de la pureza, la simplicidad y la nobleza del amor. Para otros El nacimiento de Venus simboliza la feminidad, la fertilidad y el renacimiento o primavera a partir de una concha marina, aunque esta connotación ahora se sabe que no es del todo precisa (Uffizi, 2017). Otras interpretaciones dotan a este lienzo de un contenido altamente religioso, resignificándolo como la Concha del Peregrino dentro del simbolismo del Camino de Santiago (Álvarez-Rodríguez y García-Calvo, 2012).
Lo cierto es que desde que esta diosa romana de la belleza y el amor, conocida como Venus (y como Afrodita en la mitologia griega), emerge a partir de una concha marina, a los mariscos se les asocia con los alimentos “afrodisiacos”.
Las bivalvos secretan dos conchas de tamaño regular o asimétrico, viven siempre en condiciones acuáticas y muchas especies, como las almejas y las ostras, tienen un gran valor comercial porque producen perlas naturales en su interior a partir de la nucleación de un grano o de una partícula sedimentaria. Como grupo, los bivalvos, como la gran mayoría de moluscos, constituyen un grupo de animales invertebrados con un gran éxito evolutivo y un amplio registro fósil. Entre los conchíferos actuales (aquéllos que secretan un exoesqueleto en forma de concha) se encuentran los gastrópodos, los bivalvos y los cefalópodos. Al igual que muchos otros animales, su historia evolutiva data desde hace unos 542 millones de años (Ma), cuando surgieron en un evento de diversificación biológica conocido como la “Explosión del Cámbrico”.
El proceso de biomineralización
La concha de los moluscos es un excelente ejemplo de biomineralización, es decir, del proceso celular por el cual se lleva a cabo la precipitación de minerales dentro o fuera de los seres vivos como resultado de su metabolismo en un determinado momento de su ciclo vital (Dove, De Yoreo y Weiner, 2003). La concha es secretada por el manto y está formada en más de 95% por carbonato de calcio (CaCO3 ), y dependiendo de la especie, puede precipitarse la calcita, la aragonita o algún otro polimorfo de carbonato de calcio durante la biomineralización. Este proceso implica la existencia de un programa genético que dicta el tiempo de mineralización, el espacio de precipitación de la concha y su crecimiento sucesivo (Wallace et al., 2013). En las ostras, por ejemplo, la concha se forma por la depositación secuencial de carbonato de calcio a lo largo de dos direcciones: hacia el fondo (dentro de la concha) y hacia atrás (en dirección opuesta a la charnela).
En general, la estructura de la concha presenta tres capas: una capa orgánica que sirve de protección y aislante llamada periostraco, una capa nacarada de calcita o aragonita y una capa prismática de calcita (Dove, De Yoreo y Weiner, 2003). El resto lo componen polímeros orgánicos que incluyen proteínas. La estructura macromolecular de la concha presenta grandes variaciones ya que depende de la especie y de las condiciones ambientales de crecimiento. A medida que el animal va creciendo se va añadiendo nuevo material a un templado orgánico. Tanto la calcita como la aragonita se encuentran ensamblados como una matriz orgánica en cantidades variables (figura 2).
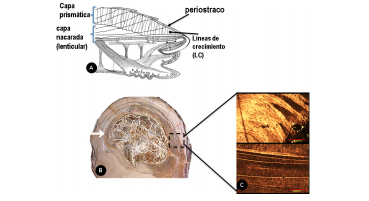
Figura 2. (A) Estructura de la concha; (B) corte transversal de Exogyra costata que muestra una zona de recristalización central (flecha); (C) sección petrográfica de laminación de la concha E. Costata
El caso de las ostras es de gran utilidad bioestratigráfica porque probablemente son un grupo monofilético. Como es un grupo abundante en facies sedimentarias que favorecen su preservación, presentan un registro fosil muy completo; especialmente durante el Cretácico, las ostras se hicieron más cosmopolitas (Wallace et al., 2013). La estructura interna de la concha de Exogyra ponderosa presenta líneas de crecimiento anuales que están constituidas por capas claras formadas durante el verano y capas delgadas negras formadas durante los meses de invierno. También durante el Cretácico las conchas de moluscos desarrollaron ornamentaciones y morfologías muy calcificadas. La concha de Exogyra ponderosa no sólo es un buen ejemplo de fauna litoral, sino también uno de los fósiles guía mejor conocidos, para el noreste mexicano y parte Texas, de los mares del Cretácico Superior (figura 3). Junto con Exogyra costata y otras conchas, se ha reportado en horizontes de la parte basal del Turoniano y del Cenomaniano Superior en varias localidades de Coahuila, Durango y Zacatecas (Böse, 1913). Estas especies eran filtradores y se enterraban a cierta altura en sustrato blando. Dichas poblaciones de moluscos filtradores epibentónicos habitaron mares calcíticos cálidos y someros durante un periodo muy cálido en la historia del planeta.
Figura 3. De manera secuencial, ejemplares fósiles de Exogyra ponderosa (A y B) y de Exogyra costata (C y D).
Exogyra ponderosa Römer se ubica taxonómicamente dentro del orden Ostreoida y dentro de la Familia Grifaeidae Vyalov 1936 (Abdelhady y Elewa, 2010). Su descripción sinóptica corresponde a una concha de una especia adulta, caracterizada por una concha gruesa y convexa en espiral que representa la valva izquierda como se aprecia en la figura 3 (con 13 cm de largo y 11cm de ancho). La concha de este ejemplar fósil conserva aún la oscura coloración de la capa nacarada original (figura 4B). En su interior se observa el crecimiento secuencial en forma de laminillas que aún se preservan en su disposición original, a diferencia de la parte lateral y central de la concha con un mayor grado de cristalización (figura 4C).
Exogyra ponderosa presenta una curvatura en espiral característica y una concha esculpida en capas. Muchos de los ejemplares aún conservan el color y la mineralogía original, especialmente en la región próxima al umbo. Desde el exterior hacia el interior de la concha, se observan zonas bien diferenciadas de la estructura de la concha: capas claras alternadas con capas oscuras en corte petrográfico (figura 4). La zona clara del periostraco, con un grosor aproximado de 1.5 cm, presenta un bandeamiento ondulante seguido de una mineralización muy calcificada de grosor constante, con contenido de aragonita en la zona interna de la concha. Las variaciones que se observan en las secciones petrográficas también sugieren temperaturas cálidas estacionales. Este patrón de bandeamiento puede ser específico y puede ser una característica variable dependiendo del ambiente y la historia de vida (Farinati et al., 2010). El análisis de las conchas permite no sólo conocer la mineralogía de la estructura interna y compararla con los ejemplares vivos, sino también inferir las condiciones ambientales del pasado.
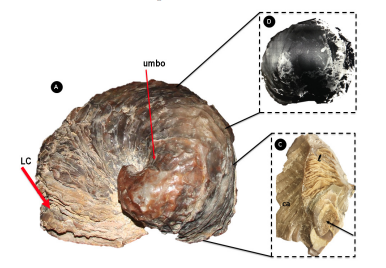
Figura 4. Secciones petrográficas de la concha de Exogyra ponderosa, las cuales muestran la alternancia estacional de laminaciones claras y oscuras.
La distribución de Exogyra ponderosa es bastante amplia, pues abarca horizontes cretácicos de los estados de Coahuila, Durango, San Luis Potosí, entre otros (Eguiluz, 2011; Franco-Rubio, 2007), y se han reportado hallazgos en diferentes formaciones, como las formaciones Olmos y Eagle Ford, pero es en la Formación San Miguel, en Coahuila, donde el registro es más abundante; en general son zonas sobreyacidas por biozonas de Exogyra costata. También se han encontrado fósiles de estas Exogyras en la Formación Cerro del Pueblo del Campaniano Tardío (Wolleben, 1977) y en la Formación Cárdenas (De la O, 2013). De igual forma se ha reportado para la Formación Cárdenas, en Durango, y en lutitas y calizas margosas de localidades del Santoniano pertenecientes a la Formación Austin, en México y en Texas, dentro de la Formación Upson que sobreyace concordantemente a la Formación Austin y subyace a la Formación San Miguel (Vejar, 1998).
Referencias
Abdelhady, A.A., y Elewa, A.M.T. (2010). Evolution of the Upper Cretaceous Oysters: Traditional Morphometrics Approach, en: A.M.T. Elewa (ed.), Morphometrics for Nonmorphometricians, Lecture Notes in Earth 157 Sciences 124, Springer-Verlag Berlin Heidelberg.
Acidini, C. (2009). Botticelli nel suo tempo. Flammarion, Mondadori Electa S.p.A., Milano, Italia, 399 pp.
Álvarez-Rodríguez, M., y García-Calvo, L. (2012). La concha del peregrino (Pecten jacobaeus), símbolo del Camino de Santiago. Ambociencias, FCByA, Universidad de León, 12-20 p.
Böse, E. (1913). Algunas faunas del Cretácico Superior de Coahuila y zonas limítrofes, Boletín Núm. 30, Instituto Geológico de México, Secretaría de Fomento, Colonización e Industria, 1-56 pp.
De la O B., F. (2013). Etude pétrographique et géochimique intégrée du charbon et de shale à gaz du bassin Sabinas et de Chihuahua au nord du Mexique: estimation des ressources en gaz méthane. PhD Tesis, l’Université de Lorraine, 395 pp.
Dove, P.M., De Yoreo, J.J., y Weiner, S. (Eds.). (2003). Biomineralization, Reviews in Mineralogy and Geochemistry, Mineralogical Society of America, Vol. 543, 84 pp.
Eguiluz de A., S. (2011). Geological synopsis of the Burgos Basin, Northeastern Mexico: petroleum production and resources, Boletín de la Sociedad Geológica Mexicana, Vol. 63, Núm. 2, p. 323-332.
Farinati, E.A., et al. (2010). Respuesta morfológica y microestructural condicionada por estrés en Crassostrea patagonica (D ́Orbigny) de la Formación Río Negro (Mio-Plioceno), Patagonia, Argentina. Revista de la Sociedad Geológica de España 23 (1-2): 9-22.
Franco-Rubio, M. (2007). Geología y paleomagnetismo de la porción centro oriental del estado de Chihuahua, México: definición del lineamiento Delicias-Mulato. Tesis doctoral, FCT, UANL, 278 pp.
Uffizi. (2017). Guide to Uffize Gallery Museum. Disponible en http://www.uffizi.org
Vasari, G. (1986). The Great Masters. Hugh Lauter Associates Inc., 387 pp.
Vejar H., M.M. (1998). Manifestaciones de hidrocarburos en ambiente deltaico de la Formación San Miguel en la subcuenca Fuentes Río Escondido, noreste de México. Tesis FCT, UANL, 115 pp.
Wallace, A.F. et al. (2013). Eukaryotic Skeletal Formation en: Knoll, A.H., Canfield, D.E., y K.O. Konhausser, Fundamentals of Geobiology, Wiley-Backwell Publ., 443 pp.
Wolleben, J. (1977). Paleontology of the Difunta Group (Upper Cretaceous- Tertiary) in northern Mexico. Journal of Paleontology, 51(2), 373-398.
*Universidad Autónoma de Nuevo León
Contacto: baicalia2012@gmail.com