La importancia de la herbivoría en el registro fósil

Patricia Velazco de León*
CIENCIA UANL / AÑO 20, No. 85, julio-septiembre 2017
El registro fósil nos permite tener una panorámica general de la evolución de los diferentes grupos de organismos y los ambientes en los que han vivido. De esta manera es posible estudiar cómo fueron las primeras formas de vida de los organismos unicelulares en diferentes localidades del Precámbrico, hasta la explosión de la diversidad de los invertebrados marinos durante el Paleozoico y la invasión de las primeras traqueofitas al ambiente continental. Esta diversidad, ligada a innovaciones morfológicas, anatómicas y reproductivas, tiene su contraparte en las extinciones en diferentes periodos que evidencian cambios climáticos, orográficos y tectónicos de los diferentes grupos en ambientes continentales y marinos.
Los insectos actores de estas líneas son la clase más diversa en nuestros días (2-100 millones, depende de los autores), tienen su registro más antiguo en el Devónico, en la localidad de Rhynie, Escocia (Penney y Jepson, 2014). Son comúnmente conservados en el registro fósil como impresiones y compresiones las extraordinarias concreciones de Mazon Creek (Baird, 1997), carbonizaciones en lignita del Cretácico, inclusiones en ámbar como las del Báltico, Chiapas y Dominicana. La evidencia indirecta de su presencia, donde quedó registrada su actividad, pertenecería a la categoría de icnofósiles. Existen diferentes propuestas de clasificación para estas actividades (Seilacher, 1953; Vialov 1975; Labandeira y Currano, 2013), aquí nos restringimos a mencionar las que registraron de manera directa sobre las plantas. La evidencia que han dejado los insectos de su interacción con las plantas está bien documentada desde el Carbonífero, por ejemplo, ovoposición endofítica en Calamites (Bethoúxs et al., 2004) la presencia de agallas en pteridospermas y Filicales, agallas de diferente forma y dimensiones, minas rectas, alimentación en el margen y esqueletización.
Herbivoría México
En México son escasos los registros de yacimientos fosilíferos de insectos, y más aún las localidades donde se haya estudiado herbivoría. Galdámez et al. (2006) y Galdámez (2006) reportan esta interacción en la Formación Atotonilco el Grande para el Plioceno, en el estado de Hidalgo; comentan la presencia de tres tipos de interacción en 34 hojas, menos de 10% de todas las hojas revisadas. Las agallas de forma redonda y los indicios de alimentación en el margen se localizaron sobre hojas de Populus tremuloides.

Figura 1 Agallas en Populus tremuloides de la Formación Atotonilco el Grande, estado de Hidalgo (Plioceno).
Debido al tamaño y forma de las agallas propusieron que insectos pertenecientes a himenópteros o dípteros pudieron haber sido los causantes. En la figura 2 se observa alimentación en el margen, en las hojas de Quercus sp. dejadas por el aparato masticador de un insecto, pero no se propuso orden. Para las minas onduladas, mencionan que pudieron haber sido realizadas por dípteros de la familia Agromizidae apoyado en su grosor, además de no ser una mina muy elaborada. El último tipo de alimentación registrado fueron los hoyos en el género Juglans que pudieron haber sido hechas por muchos ordenes de insectos con aparato bucal masticador, la flora mencionada vivió en los alrededores del paleolago de Amajac (Arellano et al., 2005).

Figura 2. Alimentación en el margen en hojas de Quercus sp. Formación Atotonilco el grande, estado de Hidalgo (Plioceno).
Remontándonos a la era Mesozoica, cuando tuvieron su clímax las gimnospermas, Lozano-Carmona (2012) registró la presencia de una galería de descortezador (figura 3).

Figura 3.Galerías de un probable descortezador de la localidad Río Ñumi en la Formación Zorrillo-tabernas indiferenciadas
En la formación Zorrillo-Taberna indiferenciadas del Jurásico Medio, Velasco de León et al. (2006) dan a conocer las evidencias de herbivoría en otra localidad de la misma formación en los géneros de Bennettitales Zamites y Otozamites donde se identificaron mordeduras en el margen de estos géneros.
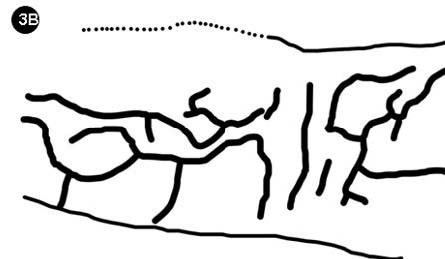
Figura 3B Reconstrucción de las galerías.

Figura 4 Daño por alimentación externa en el ápice de pina de Otozamites sp (CFZ Ot 87) de la Formación Zorrillo-Tabernas indiferenciadas. Nótese la zona oscura donde ocurrió necrosis del tejido.
En la figura 4 se puede observar un oscurecimiento que indica la zona de necrosis y la respuesta de la planta, por lo que se propone que cuando esta hoja fue comida estaba viva atada a la planta.

Figura 5 Minas sobre hojas de Otozamites hespera.
También existen minas rectas (figura 5) sobre la lámina de Otozamites hespera, se ha propuesto que este tipo de minas están asociadas a un insecto monófago, puesto que después del Triásico las interacciones internas se hacen más específicas (Scott et al., 1992; Labandeira, 2006). Continuando con el Jurásico, Díaz y Velasco de León (2017) han estudiado evidencias de interacción planta insecto en la formación Tecomazuchil sobre hojas de Bennettitales y Caytoniales, donde se reconoció la presencia de dos tipos de minas, mordeduras y agallas. Podemos concluir que la información de este tipo de interacciones nos habla de la diversidad de insectos que existieron en el Jurásico Medio viviendo sobre diferentes grupos de gimnospermas, Bennettitales, Caytoniales, etc. Para este periodo se encontraban presentes diferentes tipos de daño como minas, agallas, galerías, alimentación en el margen, que han sido relacionados con los órdenes coleóptera, lepidóptera y díptera; además de que la selección de los géneros de plantas consumidos eran más palatables. Este tipo de daño también se ha registrado para el Jurásico de Norteamérica, Australia y algunas comunidades de Europa. Por lo que se puede proponer que en el Cretácico y Neógeno, con el advenimiento de las angiospermas, los actores prevalecen (insectos) y sólo cambia el escenario, además se sustituyen las gimnospermas por las angiospermas.
*Universidad Autónoma de México
Contacto: velpaty7@gmail.com
Referencias
Arellano-Gil, J., et al. (2005). Origen y características geológicas del paleo-lago de Amajac, Hidalgo. Revista Mexicana de las Ciencias Geológicas, 22 (2):199-211.
Baird, G.C. et al. (1997). Palenvironmental setting of The Mazon Creek biota. Pp 31-51. In Shrica, C.W. and A.A. Hay (eds.). Richardson guide to the Fossil Fauna of Mazon Crek, Northeastern Illinois University; Chicago Illinoi, 308pp.
Bethoux, O., Galtier, J. y Nel, A. (2004). Earliest evidence of insect endophytic oviposition. Palaios, 19:408-318.
Carmona-Lozano, D.E. (2012). Paleoclima y flora fósil de Río Ñumi, Formación Zorrillo-Taberna indiferenciada, Oaxaca, México: México. Facultad de Estudios Superiores Zaragoza, UNAM. Tesis de licenciatura
Díaz L., R.K., y Velasco de L., M.P. (2017). Interacción planta-insecto en hoja del Jurásico Medio de las localidades Ayuquila y Chilixtlahuaca, Oaxaca. XV Congreso Nacional de Paleontologia. San Luis Potosi, México
Galdámez E., E.I. (2006). Herviboría en angiospermas fósiles de la Formación Atotonilco El Grande, Hidalgo. México: México. Facultad de Estudios Superiores Zaragoza, UNAM. Tesis de licenciatura. 36 pp.
Galdámez E., E.I., y Velasco de L., M.P. (2006). Interacción planta-insecto en fósiles de la Formación Atotonilco el Grande, Hidalgo, México. En Libro de Resúmenes IX Congreso Latinoamericano de Botánica Santo Domingo, República Dominicana, pp. 528.
Labandeira, C.C. (2006). Silurian to Triassic plant and insect clades and their associations: new data, a review, and interpretations. Arthropod Systematics & Phylogeny, 64, 53-94.
Labandeira, C.C. y Currano, E.D. (2013). The fossil record of plant-insect dynamics. Annual Review of Earth and Planetary Sciences, 41:287-311.
Lozano-Carmona, D.E. (2012). Paleoclima y flora fósil de Rio Ñumí, Formación Zorrillo-Taberna indiferenciada, Oaxaca: México. Facultad de Estudios Superiores Zaragoza, UNAM, Tesis de licenciatura, 170 pp.
Penney, D., y Jepson, J.E. (2014). Fossil insects. An introduction to palaeoentomology. Uk: Siri Scientific Press.
Scott, A.C., Stephenson, J., y Chaloner, W. (1992). Interactionand coevolution of plants and arthropods during the Palaeozoic and Mesozoic. Philosophical Transactions of the Royal Society of London, 335, 129–165.
Seilacher, A. (1953). Studien zur paläontologie: Über die methoden der palichnologie. Neus Jahrbuch für Geologie und Paläontologie. Abhandungen, 96:421-452.
Velasco de L., M.P., Flores B., M.A., Cadena G., D. (2015). Interacción planta-insecto sobre hojas de Bennettitales en la localidad Cerro el Matador del Jurásico Medio formación Otlaltepe (Puebla). Paleontología Mexicana, vol. 4 núm.2 pp1-5 ISSN: 0185-478X.
Vialov, O.S. (1975). The fossil traces of insect feedind. Paleontologichesky Sbornik, 12:147-155.