El yin y el yang de la astrogliosis reactiva

Mariana Castro Azpíroz*, Cynthia Gabriela Sámano Salazar*
CIENCIA UANL / AÑO 24, No.106, marzo-abril 2021
Por mucho tiempo, el estudio del sistema nervioso en la salud y en la enfermedad tuvo una “visión neurocéntrica”, en la que el papel estelar lo acaparaban las neuronas. Aunque las neuronas y las células gliales se describireron a principios del siglo XIX, el estudio fisiológico de la glía comenzó 50 años después. Actualmente, este punto de vista ha cambiado debido a que diversas investigaciones han revelado que la interacción glía-neurona es fundamental en procesos fisiológicos y patológicos, de tal forma que varios neurocientíficos tienen ahora una “visión gliocéntrica”.
En particular, los astrocitos son clave para que las neuronas lleven a cabo de manera apropiada sus funciones, pero también reaccionan prácticamente a todos los tipos de alteraciones patológicas que afectan la homeostasis del sistema nervioso, esto es mediante cambios morfológicos y moleculares significativos, conocidos como gliosis reactiva o astrogliosis. Sin embargo, esta respuesta se puede convertir en un arma de dos filos, ya que si bien es esencial para crear un ambiente neuroprotector y evitar la expansión del daño hacia otras regiones del sistema nervioso, también impide la adecuada regeneración neuronal en la región del daño. En este artículo resumimos y resaltamos algunas de las funciones de los astrocitos y cómo la astrogliosis reactiva, aunque sea un mecanismo de defensa del organismo, tiene su lado positivo y negativo: un yin y un yang en la respuesta al daño neuronal.
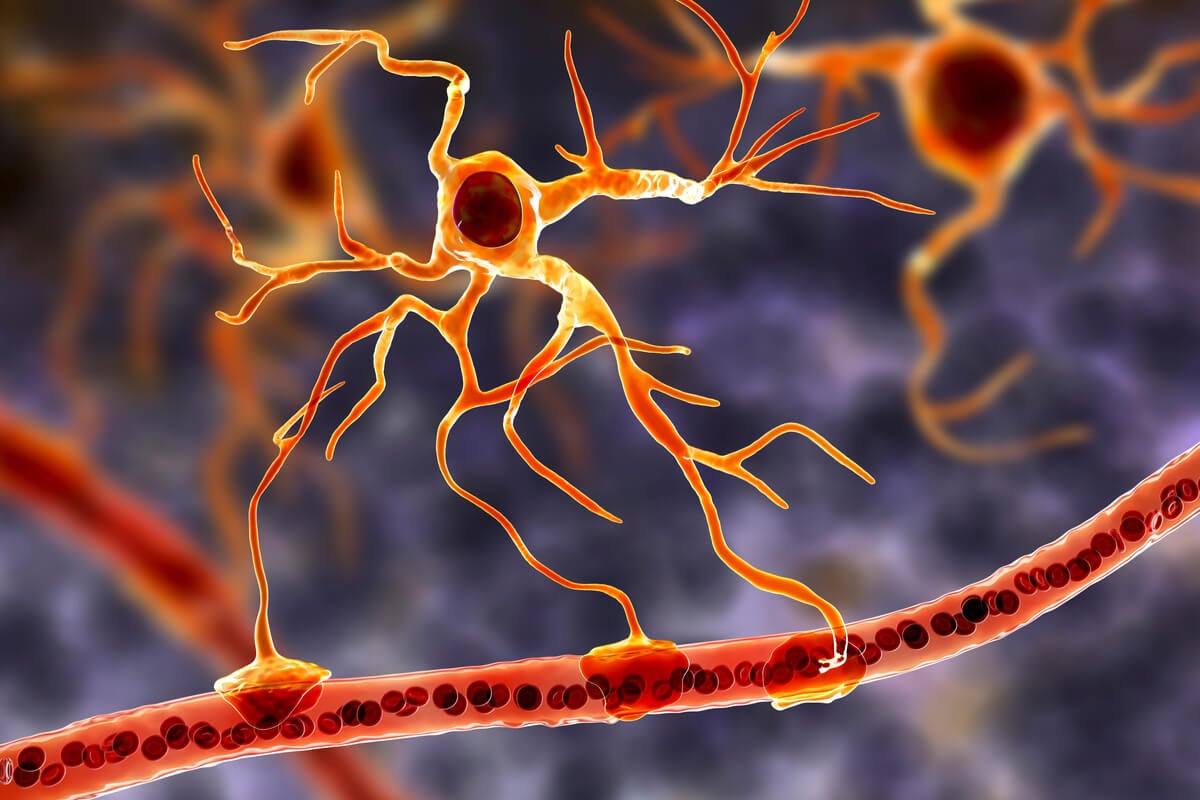
LAS “OTRAS” ESTRELLAS DEL SISTEMA NERVIOSO
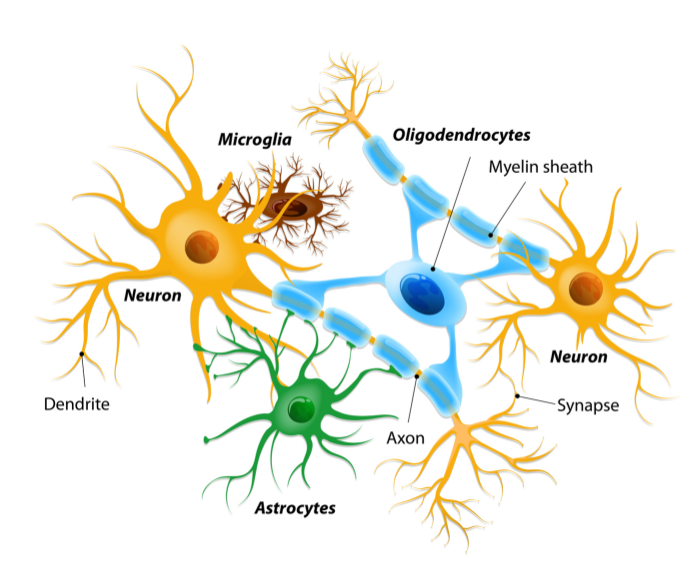
Por mucho tiempo se pensó que las neuronas eran las células nerviosas por excelencia, debido a que tienen la capacidad de conducir señales eléctricas. Sin embargo, existen otras células que también forman parte de este sistema y participan activamente en la transmisión y procesamiento de la información nerviosa. Así, el sistema nervioso está conformado por dos principales tipos de células: las neuronas y las células gliales o neuroglia. El término neuroglia fue introducido en 1858 por el médico y biólogo alemán Rudolf Virchow, quien describió la presencia de una sustancia conectiva que se formaba en el cerebro, médula espinal y en los nervios sensoriales superiores, la cual fungía como “adhesivo” en el tejido nervioso.
Ahora se sabe que se trata de la glía (del griego glia, pegamento o unión); son células que superan en número a las neuronas en una proporción de 10:1 (von Bartheld et al., 2016), proporcionan soporte a la estructura neuronal, mantienen la homeostasis del sistema nervioso, participan activamente en el procesamiento de las señales nerviosas y son defensoras del sistema nervioso durante el desarrollo y fase adulta de un organismo. Además están implicadas en algunas enfermedades neurodegenerativas y otros procesos patológicos que afectan el funcionamiento adecuado de dicho sistema (Jessen, 2004; Verkhratsky et al., 2019). La neuroglía se ha clasificado en dos principales categorías: glía del sistema nervioso periférico (SNP), que se subdivide en: 1) células de Schwann; 2) células gliales satélite; 3) células envolventes olfatorias y 4) glía entérica (Verkhratsky et al., 2019), y la glía del sistema nervioso central (SNC), que se clasifica en microglía y macroglía.
Debido a la enorme heterogeneidad celular que presenta la macroglía, se ha tratado de establecer una clasificación de acuerdo a las características estructurales y bioquímicas. Así, la macroglía constituye los siguientes tipos celulares: 1) oligodendrocitos, que pueden ser de tipo I – IV; 2) células NG2, conocidas como células progenitoras de oligondendrocitos (OPC, por sus siglas en inglés), o sinantocitos o polidendrocitos, y los 3) astrocitos o astroglia, cuyas funciones y morfología difiere en gran medida según su ubicación, subtipos y la etapa del desarrollo en la que se encuentran. De esta forma los astrocitos incluyen una amplia gama de tipos celulares: astrocitos protoplásmicos de la sustancia gris, fibrosos de la sustancia blanca, perivasculares, marginales, velatos, interlaminares, polarizados, tanicitos, pituicitos, ependimocitos, células del plexo coroideo, células epiteliales de la retina pigmentada, glía radial, glía de Müller y glía de Bergmann (Verkhratsky et al., 2019).
 El término astrocitos surgió por su apariencia estrellada debido a la gran cantidad de prolongaciones celulares que emergen de ellas; astro proviene del vocablo griego que significa estrella (Verkhratsky et al., 2019). Los astrocitos constituyen aproximadamante 30% del SNC, y tienen una variedad de funciones, entre otras: sostienen a las neuronas de forma estructural, metabólica y trófica; intervienen en la formación de la barrera hematoencefálica, ya que circundan las sinapsis, retiran el exceso de neurotransmisores y regulan la homeostasis del líquido extracelular (Sofroniew y Vinters, 2010; Verkhratsky y Nedergaard, 2018). También participan en la formación de contactos sinápticos, maduración, mantenimiento y eliminación de las sinapsis. Esto depende en gran medida de la transferencia activa de la información que va de la glía a las neuronas, a través de la fabricación y liberación de sustancias por los astrocitos, proceso conocido como gliotransmisión (Savtchouk y Volterra, 2019).
El término astrocitos surgió por su apariencia estrellada debido a la gran cantidad de prolongaciones celulares que emergen de ellas; astro proviene del vocablo griego que significa estrella (Verkhratsky et al., 2019). Los astrocitos constituyen aproximadamante 30% del SNC, y tienen una variedad de funciones, entre otras: sostienen a las neuronas de forma estructural, metabólica y trófica; intervienen en la formación de la barrera hematoencefálica, ya que circundan las sinapsis, retiran el exceso de neurotransmisores y regulan la homeostasis del líquido extracelular (Sofroniew y Vinters, 2010; Verkhratsky y Nedergaard, 2018). También participan en la formación de contactos sinápticos, maduración, mantenimiento y eliminación de las sinapsis. Esto depende en gran medida de la transferencia activa de la información que va de la glía a las neuronas, a través de la fabricación y liberación de sustancias por los astrocitos, proceso conocido como gliotransmisión (Savtchouk y Volterra, 2019).
Los astrocitos juegan un papel fundamental en la modulación de la actividad sináptica evocada por el potencial de acción (ondas de descarga eléctrica que viajan a lo largo de la membrana), en este proceso las terminales astrocíticas son fundamentales, ya que poseen receptores para otros neurotransmisores, lo que permite detectar las necesidades de las neuronas para que éstas a su vez generen una respuesta rápida ante el estímulo. En esta sinapsis tripartita interactúan un elemento presináptico, uno postsináptico y la glía asociada (Araque et al., 1999). Los astrocitos tienen en su membrana canales de sodio y potasio que pueden evocar potenciales de acción, por lo tanto son excitables, sin embargo no propagan las señales eléctricas entre ellos (Verkhratsky y Nedergaard, 2018). Pueden recibir información neuronal a través de una amplia gama de receptores membranales, para después traducirla y propagarla al circuito neuronal a través de variaciones en las concentraciones de calcio. No obstante, aún se investigan los detalles en la red de contactos integrados entre neuronas y astrocitos.
La inteligencia es otra función a la que los astrocitos se han asociado. Así, por ejemplo, el estudio histológico del cerebro de Einstein reveló que tenía un mayor número de astrocitos, los cuales, al parecer, formaban una citoarquitectura más compleja (Diamond et al., 1985). Además, se ha visto que los astrocitos, junto con los oligodendrocitos y microglia, también participan en el procesamiento y almacenamiento de recuerdos (Alberini et al., 2018). De tal forma que los astrocitos tienen un papel integral en diversas funciones del SN, pero también en la reparación de los daños en el SNC, lo cual se retomará más adelante en el texto.
UNA GAMA DE RESPUESTAS POR PARTE DE LOS ASTROCITOS ANTE EVENTOS PATOLÓGICOS
Los astrocitos tienen la capacidad de reaccionar a través de una serie de cambios morfológicos, bioquímicos, transcripcionales y funcionales ante una situación de daño en el SNC. Estos cambios constituyen la gliosis reactiva, que también se ha definido con otros términos: astrogliosis, reactividad astrocítica o astrocitosis. Existen diferentes visiones de lo que son y no son “astrocitos reactivos”, y aunque hay discrepancias para definir una única nomenclatura, se ha llegado a un acuerdo para tomar características morfofisiológicas y moleculares que diferencien los astrocitos normales de los astrocitos reactivos en sus distintos estados de reactividad. De acuerdo a la revisión que hicimos sobre el tema, en este artículo usaremos los términos astrogliosis reactiva y astrocitos reactivos para describir las formas de respuestas de los astrocitos asociados con cualquier forma de daño o enfermedad del SNC.
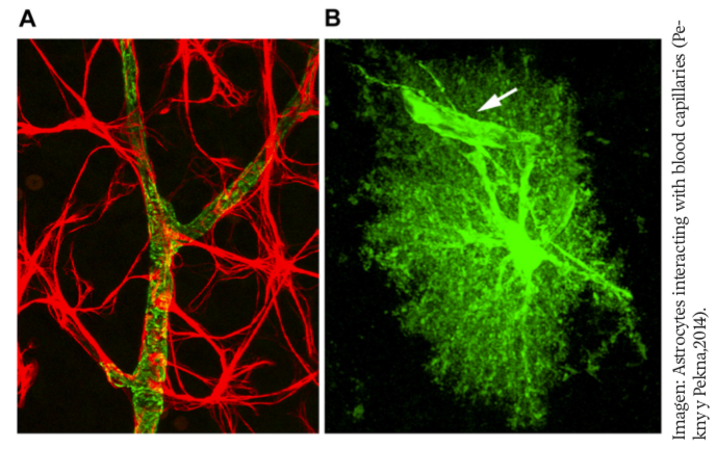
Cuando ocurre un daño o lesión en el SNC, la situación patológica se torna compleja debido a la heterogeneidad molecular y funcional de los astrocitos reactivos, donde cada subtipo de astrocitos reactivos parace ejercer una función particular en el progreso del daño (Escartin et al., 2019; Sofroniew y Vinters, 2010). Los astrocitos responden progresivamente, dependiendo de la severidad de la lesión, lo que conlleva a que la astrogliosis reactiva sea un fenómeno complejo que implica una serie de cambios secundarios finamente graduados que ocurren dependiendo del grado de daño y que están regulados, a su vez, por diversos eventos de señalización específicos (figura 1).

Figura 1. Propuesta para esquematizar los eventos fisiopatológicos que ocurren en la astrogliosis reactiva. Se muestra cronología de los astrocitos normales en estado saludable vs los mecanismos fisiopatológicos inducidos por una lesión. La activación de la astrogliosis reactiva depende del grado de la lesión. La astrogliosis puede avanzar de las fases media, severa y muy severa, y con ello se generan diversos cambios morfofisiológicos y moleculares que definen el grado de severidad de la astrogliosis.
Durante la astrogliosis hay hipertrofia de los astrocitos, en la que el citoplasma y los núcleos son agrandados y presentan una serie de procesos citoplasmáticos largos y gruesos que en condiciones fisiológicas no se observan. También hay un aumento en el número de filamentos intermedios constitutivos de los astrocitos, formados por la proteína ácida glial fibrilar (GFAP, por sus siglas en inglés), y puede incrementarse el número de células cerca de la lesión aguda. En algunos casos la proliferación puede ser discreta o ausente, pero este grado de reactividad parece depender del tipo de lesión (figura 1) (Sofroniew y Vinters, 2010). La proliferación de astrocitos puede originarse a partir de células madre multipotentes o precursores gliales todavía presentes en el SNC adulto, aunque también se ha propuesto que podrían desdiferenciarse los astrocitos maduros (Buffo et al., 2008).
Hasta ahora se han clasificado dos tipos de astrocitos reactivos, de acuerdo a (1) la gliosis isomórfica, en la que hay un patrón regular de astrocitos paralelos a los axones en degeneración. Por ejemplo, en la degeneración Walleriana, en la que la organización de las fibras gliales conserva una estructura normal. Esto también se observa en lesiones que degeneran lentamente o distantes de una lesión. Y (2) la gliosis anisomórfica, donde los astrocitos reactivos cercanos a una lesión forman una especie de malla densa sin patrón dicernible y sobreexpresan GFAP y otros marcadores de astrocitos como glutamina sintetasa (GS, una enzima esencial en los astrocitos para realizar la conversión del neurotransmisor glutamato en glutamina) y la proteína multifuncion S100β (Sofroniew y Vinters, 2010).
Un ejemplo de este tipo de gliosis son los astrocitos positivos a S100β que aumentan drásticamente en el cerebro de los pacientes con enfermedad de Alzheimer y durante otros eventos neuropatológicos, como en el síndrome de Down. Esto podría favorecer los eventos mediados por calcio en la enfermedad de Alzheimer, como la fosforilación excesiva de la proteína tau presente en marañas neurofibrilares, que finalmente podría resultar en la muerte neuronal que caracteriza a esta enfermedad (Van Eldik y Griffin, 1994).
Mediante análisis transcriptómicos se han tratado de descifrar los genes involucrados durante la astrogliosis reactiva. Así, por ejemplo, se ha visto que procesos neuroinflamatorios e isquémicos inducen dos tipos de astrocitos denominados tipo “A1” y “A2”. Los astrocitos neuroinflamatorios A1 regulan positivamente muchos genes de la cascada del complemento que son destructivos para las sinapsis, sugiriendo que los A1 tienen funciones dañinas en las células nerviosas. Mientras que los astrocitos A2 inducidos por isquemia regulan positivamente factores neurotróficos que promueven la suspervivencia y el crecimiento neuronal, por lo que se ha sugerido que los A2 tienen funciones reparadoras en el sistema nervioso.
A partir de estos hallazgos se han esclarecido algunas preguntas en relación a los diferentes estados de los astrocitos reactivos; sin embargo se desconoce si múltiples perfiles reactivos pueden ser activados en diferentes eventos patológicos (Liddelow y Barres, 2017). Lo que sí se tiene claro es que dependiendo de la severidad del daño, el proceso de gliosis puede clasificarse en tres etapas: 1) astrogliosis leve-moderada, 2) astrogliosis severa difusa y 3) astrogliosis severa con formación de tejido cicatrizante. La fase de astrogliosis leve-moderada comprende hipertrofia celular, cambios en la expresión molecular y la actividad funcional de los astrocitos. Tales cambios dependen de la gravedad del daño y ocurren después de un traumatismo leve, o en sitios distantes de una lesión más grave, cuando hay alteraciones metabólicas, infecciones leves o una reacción inflamatoria (figura2).
En la gliosis severa difusa, además de las alteraciones que ocurren en la situación leve-moderada, también hay proliferación astrocítica que provoca una reorganización duradera en la arquitectura tisular en áreas donde hay lesiones o infecciones graves. En el caso más extremo, se forma un tejido cicatrizante durante la astrogliosis severa, en los límites donde ocurren las lesiones tisulares graves, con el fin de evitar que el daño se expanda al tejido sano o restringir el avance de una infección para promover la reparación del tejido dañado. La cicatriz glial puede ser compacta, en la que los astrocitos tienen procesos densamente superpuestos, o madura si persiste por periodos prolongados y actúa como barrera no sólo para la regeneración de axones, también para las células inflamatorias, agentes infecciosos y las células que no pertenecen al SNC, de manera que protegen el tejido sano de las áreas cercanas de inflamación intensa (Sofroniew y Vinters, 2010).
Los cambios en la estructura del sistema nervioso producidos en un caso severo de astrogliosis reactiva pueden permanecer por días, meses, años e inlcuso décadas, después del inicio del daño. Esto se debe a que la astrogliosis y la formación de la cicatríz glial puede ser inducida, regulada o modulada por una amplia variedad de moléculas extracelulares liberadas por células neuronales y no-neuronales intrínsecas al SNC como microglía, oligodendrocitos, pericitos, células endoteliales y otros astrocitos, pero también células no-neurales que logran entrar al CNS (leucocitos y agentes infecciosos microbiales), los cuales liberan moléculas de señalización intercelular que van desde factores de crecimiento, citocinas, neurotransmisores, purinas, especies reactivas de oxígeno, péptido β-amiloide y reguladores de la proliferación celular.
Otros factores como: STAT3, NFκB, SOCS3, Nrf2, AMPc, Olig2, etc., participan en las vías de señalización intracelular implicadas en el aumento de GFAP, hipertrofia celular, proliferación y, en general, en la mediación de los diferentes grados de astrogliosis reactiva (Sofroniew, 2009). La astrogliosis es un proceso secundario observado durante el envejecimiento y en muchas condiciones neuropatológicas. Curiosamente, en cepas de ratones mutantes de mielina, como el ratón jimpy, la rata md y taiep, la gliosis es grave, no obstante, el crecimiento axonal no se ve afectado (Smith et al., 2013). Esto ha cuestionado el papel fisiológico de la astrogliosis con respecto a la influencia beneficiosa o perjudicial en el SNC. Los neurocientíficos han sugerido que la densa red de procesos celulares de astrocitos que pueden acumularse en el tejido cicatricial cumple funciones importantes, por un lado actúa como barrera aislando y protegiendo el tejido intacto de las lesiones, y por otro impide la regeneración axonal y tisular.
Otro tipo de alteraciones en la que están implicados los astrocitos son las astrocitopatías, las cuales se desarrollan cuando hay cambios en las funciones normales de los astrocitos en un tejido sano, estos trastornos pueden ser causados por una “astrogliosis anormal” y ser la causa principal de una astrocitopatía, disfunción neurológica y enfermedad. Las astrocitopatías fueron descritas en 2016, en Estados Unidos, y se caracterizan por la inflamación del SNC, conocida como meningoencefalitis, y la presencia de anticuerpos contra la proteína GFAP. Pueden afectar cualquier región anatómica, desde el nervio óptico hasta la médula espinal. No se conocen las causas, pero algunos casos se han asociado con diferentes tipos de cánceres, con encefalomielitis y parkinsonismo (Tomczak et al., 2019). La neuromielitis óptica es otra enfermedad autoinmune desmielinizante inflamatoria clasificada como astrocitopatía, caracterizada por la pérdida de inmunorreactividades de las proteínas GFAP y del canal permeable al agua, acuaporina 4 (AQP4, por sus siglas en inglés) que normalmente expresan los astrocitos (Lucchinetti et al., 2014). Estas enfermedades abrieron un campo de estudio, ya que aún no existen criterios uniformes de diagnóstico o un consenso para definir las astrocitopatías, y aunque se han aplicado esteroides como tratamiento, la respuesta en algunos pacientes es muy variable e incluso algunos no sobreviven.
EL LADO OSCURO DE LA ASTROGLIOSIS REACTIVA
Muchas enfermedades neurológicas tienen mecanismos en común que los astrocitos podrían contraatacar; no obstante, en muchas ocasiones los estímulos dañinos sobrepasan la capacidad de los astrocitos para generar mecanismos de neuroprotección. La astrogliosis se caracteriza por alteraciones celulares, moleculares y funcionales notables en los astrocitos reactivos (figura 2), que si no se contrarresta en la etapa postaguda o crónica temprana, después de la lesión genera consecuencias indeseadas. Por ejemplo, la desorganización de los astrocitos durante la astrogliosis reactiva puede fomentar que no se lleven a cabo correctamente las funciones de regulación de concentraciones de iones y recaptura de neurotransmisores. Además, la cicatriz glial produce altos niveles de componentes inhibidores, en particular los proteoglicanos de condroitin sulfato (CSPG, por sus siglas en inglés), que forma barreras químicas y físicas y evita el alargamiento de los axones después de una lesión en el SNC.
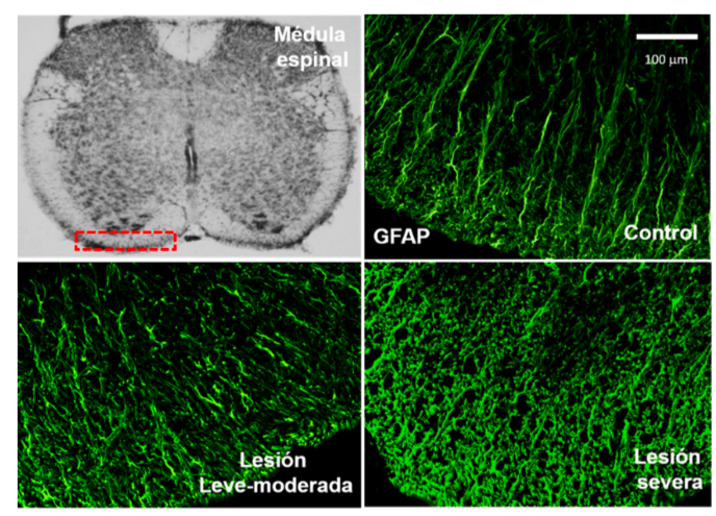
Figura 2. Ejemplo de astrogliosis reactiva que ocurre cuando se induce experimentalmente una lesión por compresión en la médula espinal. Se analizó un segmento de la sustancia blanca (recuadro punteado rojo) en tres condiciones: 1) control, donde se observan los procesos celulares de los astrocitos GFAP+ (fibras color verde); 2) lesión leve-moderada (compresión por 30 seg) produjo mayor expresión de los procesos celulares GFAP+; y 3) lesión severa (compresión por 1 min), la cual exacerbó el número de los procesos celulares GFAP+, con un evidente desarreglo en la sustancia blanca (cortes e imágenes procesadas por la Dra. Cynthia Sámano S. en el Laboratorio de Biología Celular, UAM-C).
De todas las señales inhibitorias en el entorno del SNC, los CSPG parecen ser factores clave en el blo- queo de la regeneración de axones. En varios modelos experimentales se ha demostrado que los CSPG aumentan su producción después de una lesión del SNC, y la innhibición de los CSPG que expresan las células precursoras de oligodendrocitos y astrocitos se asocia con una mejor regeneración axonal después de un traumatismo tanto en cerebro como en médula espinal (Li et al., 2016). Adicionalmente, algunos estudios transgénicos revelan el potencial que tiene la astrogliosis para exacerbar la inflamación después de una lesión traumática o incluso durante una enfermedad autoinmune. Si bien los astrocitos reactivos están presentes en el tejido del SNC dañado o degenerado, se les han atribuido directamente a la generación de efectos negativos, sin embargo existen puntos de vista opuestos en relación a si los astrocitos reactivos se pudieran considerar “amigos o enemigos”.
EL LADO AMISTOSO DE LA ASTROGLIOSIS REACTIVA
Por mucho tiempo, la astrogliosis fue considerada como una reacción fundamentalmente dañina para la recuperación funcional. Esta noción ha sido refutada por los estudios experimentales que han mostrado que en respuesta a la mayoría, si no es que a todas las agresiones del SNC, la astrogliosis inicia como un proceso adaptativo y beneficioso dirigido a la reparación de heridas y preservación de la función neurológica. Sin embargo, es importante comprender que también genera efectos negativos que son regulados por mecanismos de señalización específicos que dependerán del grado y tipo de daño en el SNC. Curiosamente, diversos estudios experimentales han mostrado que la astrogliosis es un proceso necesario, ya que su inhibición aumenta la inflamación, el daño tisular y limita la recuperación funcional después de una lesión traumática en el SNC o una lesión isquémica. Otros estudios han mostrado que el control del pH que ejercen los astrocitos puede reducir la excitotoxicidad del glutamato durante la isquemia (Pekny y Pekna, 2014).
Entre otros beneficios de la astrogliosis reactiva, reduce la infiltración de leucocitos y promueve la reparación de la barrera hemato-encefálica, limita la neurodegeneración y ralentiza la progresión de enfermedades neurodegenerativas, también limita la pérdida neuronal en un accidente cerebrovascular isquémico y neurotrauma, debido a que contrarresta el estrés hiposmótico y remueve el exceso de especies reactivas de oxígeno (Sofroniew, 2009; Sofroniew y Vinters, 2010). Pero quizá el efecto que se considera más perjudicial de la astrogliosis es la inhibición de la regeneración del axón por las cicatrices astrogiales, después del daño en el SNC. No obstante, este efecto perjudicial debe considerarse también en el contexto de las funciones protectoras esenciales que desempeñan las cicatrices de los astrocitos, porque restringen la inflamación y promueven la protección del tejido sano adyacente a las lesiones (Sofroniew, 2009; Sofroniew y Vinters, 2010; Pekny y Pekna, 2014).
De hecho, un gran hallazgo fue que la presencia de la cicatriz astroglial, cuando se combina con un gel que contiene factores de crecimiento, promueve el recrecimiento de axones después de inducir daño en la médula espinal en ratones (Liddelow y Barres, 2016). Estos datos han hecho que se cuestione si prevenir o eliminar las cicatrices de los astrocitos hace más daño que bien.
De todas las señales inhibitorias en el entorno del SNC, los CSPG parecen ser factores clave en el bloqueo de la regeneración de axones. En varios modelos experimentales se ha demostrado que los CSPG aumentan su producción después de una lesión del SNC, y la innhibición de los CSPG que expresan las células precursoras de oligodendrocitos.
CONCLUSIONES
La astrogliosis reactiva es un proceso adaptativo y conservado en la evolución; sin embargo, es un arma de dos filos: si bien es un mecanismo de defensa que cumple funciones esenciales de reparación y protección para minimizar el daño inicial en los tejidos después de las lesiones del SNC, en determinadas situaciones puede mostrar efectos dañinos, lo cual dependerá de diversos factores y situaciones para que una u otra condición se desarrolle. Finalmente, conocer y estudiar a profundidad este mecanismo conlleva a explorar la posibilidad de encontrar posibles blancos terapéuticos cuando se presenta un proceso de astrogliosis reactiva en diferentes situaciones neuropatológicas.
*Universidad Autónoma Metropolitana,
Unidad Cuajimalpa (UAM-C).
Contacto: csamano@cua.uam.mx
REFERENCIAS
Alberini, C.M., Cruz, E., Descalzi, G., et al. (2018). Astrocyte glycogen and lactate: New insights into learning and memory mechanisms. Glia. 66(6):124-1262.https://doi.org/10.1002/ glia.23250
Araque, A., Parpura, V., Sanzgiri, R.P., et al. (1999). Tripartite synapses: glia, the unacknowledged partner. Trends Neurosci. 22(5):208-15.
Buffo, A., Rite, I., Tripathi, P., et al. (2008). Origin and progeny of reactive gliosis: A source of multipotent cells in the injured brain. Proceedings of the National Academy of Sciences of the United States of America. 105(9):3581-3586. https://doi. org/10.1073/pnas.0709002105
Diamond, M.C., Scheibel, A.B., Murphy, G M.,Jr, et al. (1985). On the brain of a scientist: Albert Einstein. Experimental neurology. 88(1):198- 204. https://doi.org/10.1016/0014- 4886(85)90123-2
Escartin, C., Guillemaud, O., y Carrillo-de Sauvage, M.A. (2019). Questions and (some) answers on reactive astrocytes. Glia. 67(12):2221-2247. https:// doi.org/10.1002/glia.23687
Jessen, K.R. (2008). Glial cells. The international journal of biochemestry & cell biology. 36(10):1861-1867. https:// doi.org/10.1016/j.biocel.2004.02.023
Liddelow, S.A., y Barres, B.A. (2016). Regeneration: Not everything is scary aboutaglialscar. Nature. 532(7598):182- 183. https://doi.org/10.1038/nature17318
Liddelow, S.A., y Barres, B.A. (2017). Reactive Astrocytes: Production, Function, and Therapeutic Potential. Immunity. 46(6):957-967. https://doi. org/10.1016/j.immuni.2017.06.006
Li, K., Li, J., Zheng, J., et al. (2019). Reactive Astrocytes in Neurodegenerative Diseases. Aging and disease. 10(3):664-675.https://doi.org/10.14336/ AD.2018.0720
Li, G., Cao, Y., Shen, F., et al. (2016). Mdivi-1 Inhibits Astrocyte Activation and Astroglial Scar Formation and Enhances Axonal Regeneration after Spinal Cord Injury in Rats. Frontiers in cellular neuroscience. 10:241. ht- tps://doi.org/10.3389/fncel.2016.00241
Lucchinetti, C.F., Guo, Y., Popescu, B.F., et al. (2014). The pathology of an autoimmune astrocytopathy: lessons learned from neuromyelitis optica. Brain pathology (Zurich, Switzerland). 24(1):83-97. https://doi. org/10.1111/bpa.12099
Mi sistema inmune. (2017). Neurons and neuroglial cells. Disponible en: https://www.misistemainmune.es/ la-microglia-en-el-sistema-nervioso-central/
Pekny, M., y Pekna, M. (2014). Astrocyte reactivity and reactive astrogliosis: costs and benefits. Physiological reviews. 94(4):1077-1098. https://doi. org/10.1152/physrev.00041.2013
avtchouk, I., y Volterra, A. (2018). Gliotransmission: Beyond Black-and-White. The Journal of neuroscience: the official journal of the Society for Neuroscience. 38(1):14-25. https://doi. org/10.1523/JNEUROSCI.0017-17.2017
Smith, C.M., Cooksey, E., y Duncan, I.D. (2013). Myelin loss does not lead to axonal degeneration in a long-lived model of chronic demyelination. The Journal of neuroscience: the official journal of the Society for Neuroscience. 33(6):2718-2727. https://doi. org/10.1523/JNEUROSCI.4627-12.2013
Sofroniew, M.V. (2009). Molecular dissection of reactive astrogliosis and glial scar formation. Trends in neurosciences. 32(12):638-647. https://doi. org/10.1016/j.tins.2009.08.002
Sofroniew, M.V., y Vinters, H.V. (2010). Astrocytes: biology and pathology. Acta neuropathologica. 119(1):7-35. https://doi.org/10.1007/s00401-009- 0619-8
Tomczak, A., Su, E., Tugizova, M., et al. (2019). A case of GFAP-astroglial autoimmunity presenting with reversible parkinsonism. Multiple sclerosis and related disorders. 39:101900. Advance online publication. https://doi. org/10.1016/j.msard.2019.101900
Van Eldik, L.J., y Griffin, W.S. (1994). S100 beta expression in Alzheimer’s disease: relation to neuropathology in brain regions. Biochimica et biophysica acta. 1223(3):398-403.
Verkhratsky, A., y Nedergaard, M. (2018). Physiology of Astroglia. Physiological reviews. 98(1):239-389. https:// doi.org/10.1152/physrev.00042.2016
Verkhratsky, A., Ho, M.S., Zorec, R., et al. (2019). The Concept of Neuroglia. Advances in experimental medicine and biology. 1175:1-13. https://doi. org/10.1007/978-981-13-9913-8_1
von Bartheld, C.S., Bahney, J., y Herculano-Houzel, S. (2016). The search for true numbers of neurons and glial cells in the human brain: A review of 150 years of cell counting. The Journal of comparative neurology. 524(18):3865-3895.https://doi. org/10.1002/cne.24040