Regulación de genes que afectan la biosíntesis de compuestos de azufre en cerveza

JESSICA G. HERRERA GAMBOA*, CLARA S. LEAL GUERRA*, ESMERALDA PÉREZ ORTEGA*, LUIS C. DAMAS
BUENROSTRO**, JUAN C. CABADA**, LUIS J. GALÁN WONG*, BENITO PEREYRA ALFÉREZ*
CIENCIA UANL / AÑO 17, No. 67, MAYO-JUNIO 2014
La producción de cerveza continúa siendo un arte aun después de su industrialización. Mantener los parámetros de calidad y aceptación sin variar el sabor único de cada marca es el mayor reto para la industria cervecera. (1)
El mosto es el líquido que provee los nutrientes asimilados por la levadura durante la fermentación alcohólica. En este proceso, el etanol y el dióxido de carbono son los principales productos. Sin embargo, junto a éstos se generan subproductos, los cuales tienen efecto sobre la calidad sensorial de la cerveza. Entre otros, destacan los compuestos volátiles de azufre (CVA): el sulfuro de hidrógeno (H2S), el dióxido de azufre (SO2) y el dimetil sulfuro (DMS).
Los CVA son de los parámetros más difíciles de controlar, ya que su composición y concentración varía en función de los componentes del mosto, condiciones de manejo de la levadura, variables de fermentación y tipo de cepa. (2) La formación de H2S está relacionada con el metabolismo de la levadura. El ión HS- es un intermediario metabólico necesario en la reducción de sulfato o sulfitos para la biosíntesis de compuestos orgánicos azufrados como metionina, cisteína o S-adenosilmetionina; asimismo, tripéptidos como el tiol glutatión, los cuales son requeridos para la síntesis de proteínas. La concentración de algunos de estos aminoácidos afecta la formación de sulfuro de hidrógeno (H2S) y dióxido de azufre (SO2) durante la fermentación. (2,3)
Además de las características genéticas de la levadura que determinarán la forma en que metabolizarán los aminoácidos, algunas prácticas cerveceras pueden favorecer la producción de CVA: la maceración de la malta, la nutrición de la levadura, las altas temperaturas de fermentación, la autolisis de levadura en el medio, la pobre eliminación del gas dióxido de carbono, entre otras. (4)
Debido a su contribución aromática, la formación de estos compuestos en bebidas fermentadas ha sido sujeto de interés, ya que puede modificar las propiedades sensoriales del producto, dependiendo de su identidad y concentración. (5)
La levadura requiere la activación de ciertos genes que codifican las enzimas necesarias para la incorporación del ión HS- a distintas vías metabólicas, tal es el caso de los genes relacionados con la asimilación de sulfatos (MET3, MET14, MET16), asimilación de sulfitos (MET5, MET10, MET25, MET2, STR4, STR1), o bien, síntesis de algunos aminoácidos azufrados (MET6, MET13, SAM1, SAM2). (6)
Se ha demostrado que el tipo de cepa de levadura, y por consiguiente su carga genética, es una variable importante en la producción de H2S, ya que responden de diferente forma bajo ciertas condiciones medioambientales y fisiológicas. (7 )
Por consiguiente, existe la necesidad de un mayor conocimiento sobre la expresión de genes relacionados con compuestos de azufre bajo diferentes condiciones de operación. Más aún, la caracterización de los niveles de expresión de estos genes ayudaría en gran medida a optimizar el proceso cervecero, permitiendo predecir o regular la producción de compuestos que impacten las características finales de la cerveza.
MATERIALES Y MÉTODOS
Biológicos
Las cepas de levadura usadas en este estudio fueron Saccharomyces sp. 790 y 820, tipo “Lager”, proporcionados por Cervecería Cuauhtémoc Moctezuma (CCM).
Mosto
Los mostos se prepararon en la planta experimental de CCM. Se utilizaron dos formulaciones con diferentes porcentajes de malta y adjuntos. Para el mosto 1 se utilizó una concentración de malta menor a 50%; y para el mosto 2, mayor a 50%. Cada mosto tuvo una concentración de extracto fermentable de aproximadamente 17 grados plato (°P).
Concentración de nutrientes en mostos
Los mostos fueron muestreados asépticamente para la determinación de nitrógeno libre asimilable (FAN), siguiendo el protocolo Wort-12 de la ASBC.
Fermentaciones
En condiciones asépticas, los mostos fueron oxigenados a una concentración de 1 ppm/°P, e inoculados con 1.2X106 cel/ml por °P, para todas las condiciones. Las fermentaciones se llevaron a cabo en probetas estériles de 2 L, por siete días, a una temperatura inicial de 11°C ± 1°C el día 1, y a 16°C ± 1°C los días posteriores. La levadura fue cosechada y se analizó la viabilidad celular por la técnica de microcultivo. Para el análisis de cuantificación por PCR cuantitativa (qPCR), se utilizó un diseño factorial de dos niveles con dos factores (cepa y mosto), por duplicado, y se generaron así ocho fermentaciones en total.
Análisis transcripcional global
Al segundo día de fermentación se extrajo el ARN total de las células en suspensión de ambas cepas con el protocolo de extracción de Schmitt et al. (8) El análisis del transcriptoma se realizó mediante microarreglos de ADN con las matrices de Saccharomyces cerevisiae YOMHW_01 preparadas por el Dr. Jorge Ramírez, del Instituto de Fisiología Celular de la Universidad Nacional Autónoma de México. Los resultados fueron analizados mediante el software
GenArise con un Z score de ± 1.5. En nuestro análisis, comparamos la expresión genética con el mosto 2 como condición normal, y el mosto 1 como condición prueba.
RESULTADOS
Viabilidad de la levadura
Los resultados de la viabilidad de las cepas mostraron que este factor se mantuvo por encima de 90%, lo cual significa que nuestras cepas se mantuvieron viables durante todo el proceso.
Concentración de nutrientes en mostos
Dos tipos de mostos fueron formulados con diferentes concentraciones de malta para obtener diferentes perfiles de nutrientes. La concentración de FAN en los mostos demostró una relación de acuerdo a la concentración de malta. De esta manera, el mosto 1 tuvo una concentración de 133 ppm, mientras que en el mosto 2 fue de 171 ppm.
Análisis de expresión global por medio de microarreglos
El análisis de la expresión genética se realizó con el programa GenArise, tomando como criterio de discriminación un valor de “Z score” >1.5. De esta manera, los genes presentaron cambios en sus niveles de expresión con valores mayores a 1.5 (sobreexpresados) y menores a -1.5 (genes reprimidos) (figura 1). La tabla I muestra un resumen de los resultados de expresión de algunos genes relacionados con la generación CVA, al usar el mosto 2 como condición normal y el mosto 1 como prueba. En este análisis se demuestra la diferencia entre ambas cepas. De esta manera, la cepa 820 mostró seis genes por debajo de –1.5 (reprimidos) y uno mayor a 1.5 (sobreexpresados). La cepa 790 demostró sólo un gen reprimido y uno sobreexpresado. Ambas cepas sobreexpresan un gen en común y reprimen otro. La ubicación de cada gen en su ruta metabólica se muestra en la figura 2.
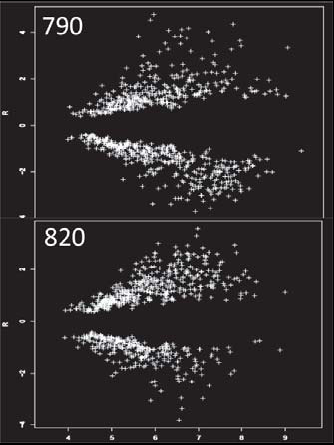
Fig. 1. Imágenes del análisis del software GenArise, tomando como criterio un
valor “Z score” de +1.5 para las cepas 790 y 820.
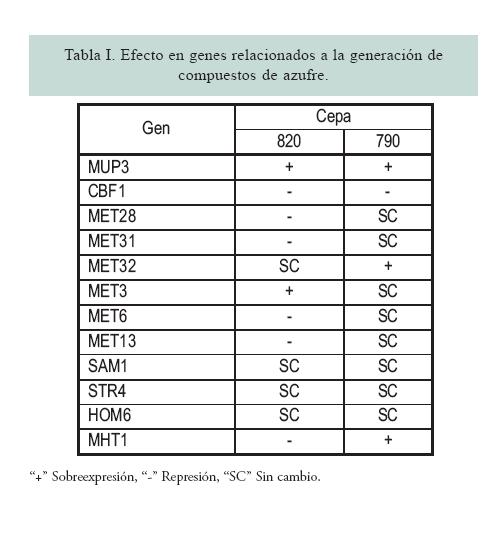
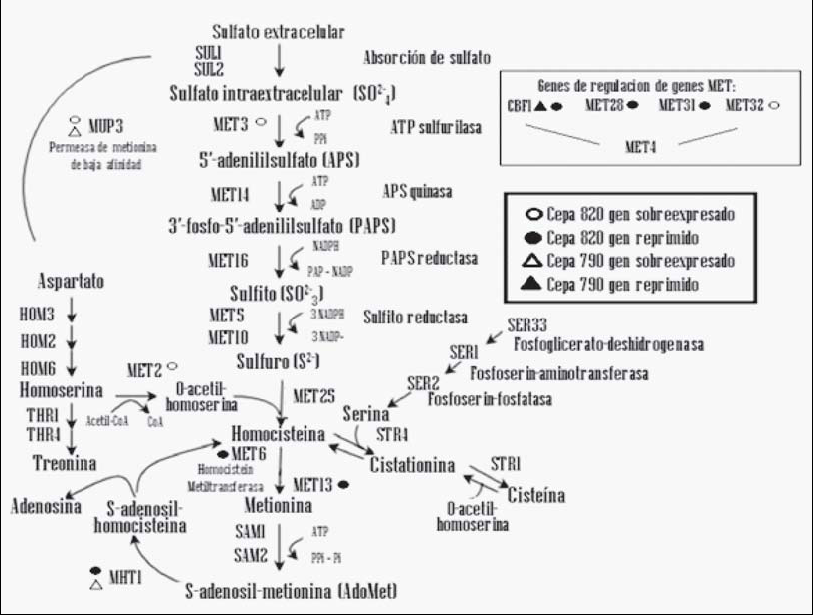
Fig. 2. Ubicación de genes seleccionados por microarreglos, en la ruta metabólica de la asimilación de sulfatos, reducción de sulfito y sulfuro y la incorporación de
aminoácidos azufrados. Modificado de Thomas and Surdin-Kerj.9
DISCUSIÓN
La producción de compuestos de azufre de dos levaduras cerveceras tipo lager, 790 y 820, se evaluó a nivel transcripcional, analizando las variables cepa y mosto, al segundo día de fermentación. El análisis del transcriptoma demostró diferencias importantes en la regulación de la expresión genética. Los resultados obtenidos indican que ambas cepas sobreexpresan el gen MUP3, el cual codifica una permeasa de metionina de baja afinidad,10 debido a que aún está presente algo de este aminoácido en el medio, el cual se consume en las primeras 48 hrs de fermentación. (11) La cepa 820 introduce mayor cantidad de sulfato a la ruta, e introduce al esqueleto carbonado para la formación de homocisteína (MET3), (12) sin ser capaz de sobreexpresar genes que eviten la acumulación de H2S (MET28, MET6 y SAM1).13-15 En el mosto 1, al tener menor nivel de FAN, insuficientes precursores de metionina y cisteína (O-acetil serina y O-acetil homoserina) estuvieron disponibles, y el H2S libre se acumuló en la célula para, finalmente, liberarse al medio. En presencia de mosto 2 sobreexpresa la vía de transulfuración y la vía de remetilación (MHT1). La capa 790 sobreexpresa genes que facilitan la incorporación del H2S para la formación de aminoácidos, sobreexpresando el gen MHT1, el cual tiene la función de mantener en equilibrio la relación entre metionina y S-adenosilmetionina (donador de grupos metilo en reacciones de transmetilación). Esto sugiere que la cepa 790 tiene la capacidad de convertir con mayor intensidad el S-adenosilhomocisteína en homocisteína y éste, por acción de otras enzimas, en metionina.16 Además, sugiere que la cepa 790 tiene la capacidad de adaptarse al tipo de mosto, y evita la acumulación de sulfuro por medio de la vía de transulfuración y remetilación; ajusta su maquinaria de expresión genética para evitar la acumulación de H2S en mostos con distintos niveles de nutrientes nitrogenados.
CONCLUSIONES
En este estudio se demostró que la producción de compuestos azufrados depende de la constitución genética de la cepa y su interacción con los nutrientes nitrogenados presentes en mostos. El comportamiento de las cepas fue diferente frente a cada tipo de mosto: la cepa 820, en mostos con baja concentración de compuestos nitrogenados, favorece la incorporación de sulfuro a esqueletos carbonatados, mientras que la cepa 790 favorece la incorporación del sulfuro hacia la transulfuración o remetilación en cualquiera de los mostos, al compensar variaciones en los compuestos nitrogenados. Este estudio sugiere que, a nivel de producción industrial, la cepa 820 tendrá un mejor desempeño, si se fermenta en mostos ricos en FAN, y la cepa 790 se puede utilizar en mostos bajos en FAN con el fin de optimizar costos.
RESUMEN
Durante la fermentación alcohólica, la levadura producirá submetabolitos que alteran las características de la cerveza. Los compuestos volátiles de azufre (CVA) son generados por la activación de genes involucrados en el metabolismo de asimilación de sulfatos, sulfitos y síntesis de aminoácidos. Identificamos los genes que participan en la producción de CVA y su respuesta bajo distintas condiciones de proceso. Utilizamos dos cepas de levadura sometidas a diferentes tipos de mosto y evaluamos su respuesta genética. Los resultados mostraron que la producción de CVA depende de la constitución genética de la cepa y su interacción con el mosto.
Palabras clave:
Compuestos volátiles de azufre, Cerveza, Fermentación.
ABSTRACT
During brewing process, the yeast will produce secondary metabolites altering the characteristics of the beer. Volatile sulfur compounds (CVA) are generated by activation of genes involved in the metabolism of assimilation of sulfates, sulfites, and amino acid synthesis. We identified genes involved in the production of VCA and its response under different process conditions. We used two yeast strains fermenting different types of worts, and we evaluated their genetic response. The results showed that the production of CVA depends on the genetic constitution of the strain and its interaction with the wort.
Keywords:
Brewing process, Sulfur compounds, Beer.
REFERENCIAS
1. Vermeulen C., Gijs L. and Collin S. 2005. Sensorial contribution and formation pathways of thiols in foods: A review. Food Rev. Inter. 21: 69-137.
2. Duan W., Roddick F. A., Higgins V. J. and Rogers P. J. 2004. A parallel analysis of H2S and SO2 formation by brewing yeast in response to sulfur-containing amino acids and ammonium ions. J. Am. Soc. Brew. Chem. 62: 35-41.
3. Heggart H. M., Margaritis A., Pikington H., Stewart R. J., Dowhanick T. M. and Russell I. 1999. Factors affecting yeast viability and vitality characteristics: a review. MBAA Tec. Quart. 36: 383-406.
4. Hardwik W. 1995. Quality control. En: Handbook of Brewing. Hardwick, W. A. (editor). Marcel Dekker Inc., USA, pp. 587.
5. Laundaud S., Helink S. and Bonnarme P. 2008. Formation of volatile sulfur compounds and metabolism of methionine and other sulfur compounds in fermented foods. Appl. Microbiol. Biotechnol. 77: 1191-1205.
6. Thomas D., Becker A. and Surdin-Kerjan Y. 2000. Reverse methionine biosynthesis from S-Adenosylmethionine in eukaryotic cells. J. Biol. Chem. 275: 40718–40724.
7. Linderholm A. L., Carrie L., Kumar G., Hong Y. and Bisson L. F. 2008. Identification of genes affecting hydrogen sulfide formation in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 74: 1418–1427.
8. Scmitt M.E., Brown T. A. and Trumpower B. L. 1990. A rapid and simple method for preparation of RNA from Saccharomyces cerevisiae. Nucleic Acids Res. 18:10.
9. Thomas D. and Surdin-Kerjan Y. 1997. Metabolism of sulfur amino acids in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 61:503-532.
10. Isnard A. D., Thomas D. and Surdin-Kerjan Y. 1996. The study of methionine uptake in Saccharomyces cerevisiae reveals a new family of amino acid permeases. J. Mol. Biol. 262: 473–484.
11. Lekkas C., Steward G., Hill A., Taidi B. and Hudgson J. 2005. The importance of free amino nitrogen in wort and beer. Tech. Q. Master. Brew. Assoc. Am. 42: 113- 116.
12. Korch C., Mountain H. A., Gyllang H., Winge M. and Brehmer P. 1991. A mechanism for sulfite production in beer and how to increase sulfite levels by recombinant genetics. En: Proceedings of the European Brewing Convention Congress. 201–208.
13. Mountain H. A., Bystro ˜m A. S., Tang Larsen J. and Korch C. 1991. Four major transcriptional responses in the methionine/threonine biosynthetic pathway of Saccharomyces cerevisiae. Yeast 7: 781–803.
14. Thomas D. and Surdin-Kerjan Y. 1991. The synthesis of the two S- adenosyl-methionine synthetases is differently regulated in Saccharomyces cerevisiae. Mol. Gen. Genet. 226: 224–232.
15. Cherest H., Davidian J. C., Thomas D., Benes V., Ansorge W. and Surdin-Kerjan Y. 1997. Molecular characterization of two high affinity sulfate transporters in Saccharomyces cerevisiae. Genetics 145: 627–635.
16. Thomas D., Becker A. and Surdin-Kerjan Y. 2000. Reverse methionine biosynthesis from S-Adenosylmethionine in eukaryotic cells. J. Biol. Chem. 275: 40718–40724.
Recibido: 19 de agosto de 2013
Aceptado:19 de septiembre de 2013