Control de la virulencia de cepas patógenas de Escherichia coli por antimicrobianos naturales
Alam García-Heredia*, Santos García*, Ángel Merino-Mascorro*, Norma Heredia*
CIENCIA UANL / AÑO 21, No. 87 enero-febrero 2018
RESUMEN
Varios grupos de Escherichia coli pueden causar enfermedades con alta mortalidad en el humano, facilitando esta capacidad por mecanismos bacterianos que permiten su colonización y persistencia. Se han desarrollado tratamientos para su control, sin embargo, debido a su eficacia incompleta y a tendencias actuales, se está contemplando el uso de compuestos naturales antimicrobianos. En este trabajo se estudió la actividad del orégano mexicano, palo de Brasil y sus principales componentes antimicrobianos sobre el crecimiento, movilidad swarming, producción de biopelículas y expresión de genes de virulencia de E. coli enterohemorrágica, E. coli enteroagregativa y E. coli shiga-toxigénica EAEC O104:H4.
Palabras clave: E. coli, biopelícula, swarming, factores de virulencia E. coli O104:H4.
ABSTRACT
Several groups of Escherichia coli may cause diseases with high mortality in human beings faciliating this capacity by baterial mechanisms which allow its settlement persistence. It’s been developed treatments for its management, nevertheless due to its incomplete effectiveness and current tendencies, the use of natural antimicrobials compoundsare being contemplated. This work studies the activity of mexican oregano, nrazilian rosewood and its main antimicrobials compounds about growth, swarming movement, biofilm production and virulence genes expression of enterohemorrhagic E. coli, enteroaggregative E. coli and Shiga-toxin E. coli EAEC O104:H4.
Keywords: E. coli, biofilm, swarming, virulence factors E.coli 0104:H4.
Las enfermedades trasmitidas por el consumo de alimentos contaminados representan un aspecto de interés global. Cada año la cepa E. coli O157:H7 de patotipo enterohemorrágico (EHEC) es responsable de 3,268 hospitalizaciones tan sólo en los Estados Unidos, algunos de estos desencadenando complicaciones severas y alrededor de 0.5% de los casos resultan en la muerte del paciente (Fouladkhah et al., 2013). La patogénesis del serotipo O157:H7 es debida a la producción de factores de virulencia específicos como la proteína intimina, la cual media la adherencia de la bacteria al intestino (Melton-Celsa, 2014), y la toxina Shiga (Law, 2000) que inhibe la síntesis de proteínas desencadenando la muerte celular. Adicionalmente, algunos mecanismos fisiológicos como la movilidad tipo swarming y la formación de biopelícula ayudan a la colonización y persistencia en diferentes ambientes (García-Heredia et al., 2013).
Recientemente otro serotipo de E. coli, el O104:H4 (EAEC O104:H4), ha emergido como un problema serio de salud pública. Esta cepa pertenece al patotipo enteroagregativo (EAEC), capaz de adherirse a las células epiteliales por medio de una fimbria de adherencia llamada AAF (Boisen et al., 2014); sin embargo, esta cepa presenta la particularidad de también producir Shiga to- xina (Stx2), que es característica del patotipo EHEC. Esta combinación de propiedades virulentas, aunado al amplio espectro de resistencia a antibióticos que presenta, produjo cuadros severos en 2011, resultando en más de 800 casos de síndrome urémico hemolítico y 53 decesos (Guy et al., 2013).
La terapia con varios antibióticos no siempre está recomendada en infecciones producidas por E. coli (Bielaszewska et al., 2012), por lo que prevenir o minimizar la contaminación por esta bacteria es la primera opción para reducir el riesgo de enfermedades en la población. Por la variabilidad de flora vegetal existente, la inmensidad de compuestos asociados y la aceptación pública de los productos naturales, resulta importante determinar la factibilidad de uso de productos naturales para controlar de manera efectiva a estas bacterias. Los extractos de la planta Haematoxylon brassiletto (Hb), comúnmente conocida como palo de Brasil, han mostrado actividad contra el crecimiento, la producción de la toxina Shiga y la adhesión de EHEC O157:H7 a células HeLa (Heredia et al., 2005), en donde se ha establecido que los neoflavonoides (brasilina y hematoxilina) tienen gran actividad antimicrobiana (Rivero-Ruiz, 2008). De igual forma, los extractos de la planta Lippia graveolens Kunth (synon. L. berlandieri Schauer), conocida como orégano mexicano, han mostrado inhibición del crecimiento contra bacterias y hongos de interés alimentario (Ávila-Sosa et al., 2010). Las propiedades antimicrobianas de este extracto han sido atribuidas principalmente a los aceites esenciales timol y carvacrol (Quintanilla-Licea et al., 2014).
Aunque algunos de estos compuestos de plantas han mostrado tener actividad anti E. coli O157:H7 (Orue et al., 2013; Kim et al., 2011), la variabilidad fisiológica entre cepas impide predecir su efecto en otras, y no existen estudios sobre el efecto en cepas EAEC, incluyendo a la altamente patógena EAEC O104:H4.
METODOLOGÍA
Antimicrobianos naturales
Las hojas de orégano y de corteza de Hb se obtuvieron de mercados locales y se realizaron extractos etanólicos (100 g de material vegetal molido y seco con 500 ml de etanol a 96%). Los extractos se maceraron a temperatura ambiente por 24 a 48 h y se filtraron con papel Whatman no. 1. Después, las muestras se depositaron en platos de vidrio para evaporar el etanol y los extractos secos se resuspendieron en una pequeña cantidad de etanol a 96% y almacenaron a 4°C en frascos ámbar. Una alícuota se utilizó para determinar el peso seco.
Los compuestos carvacrol y brasilina se adquirieron comercialmente. El antibiótico rifaximina y el citral se usaron como controles y se adquirieron comercialmente. La brasilina, carvacrol y citral fueron disueltos en agua destilada (Somolinos et al., 2009), en tanto que la rifaximina se disolvió en etanol a 96%. Todas las soluciones se almacenaron a 4°C en frascos ámbar hasta su uso.
Cepas bacterianas
En este trabajo se utilizaron la cepa EHEC serotipo O157:H7 (ATCC 43894), EAEC 042 (serotipo O44:H18) y la cepa EAEC serotipo O104:H4. Una alícuota se transfirió a tubos con 5 ml de caldo Mueller Hinton (MH,) o caldo Luria Bertani (LB) y se incubaron a 37°C/24 h.
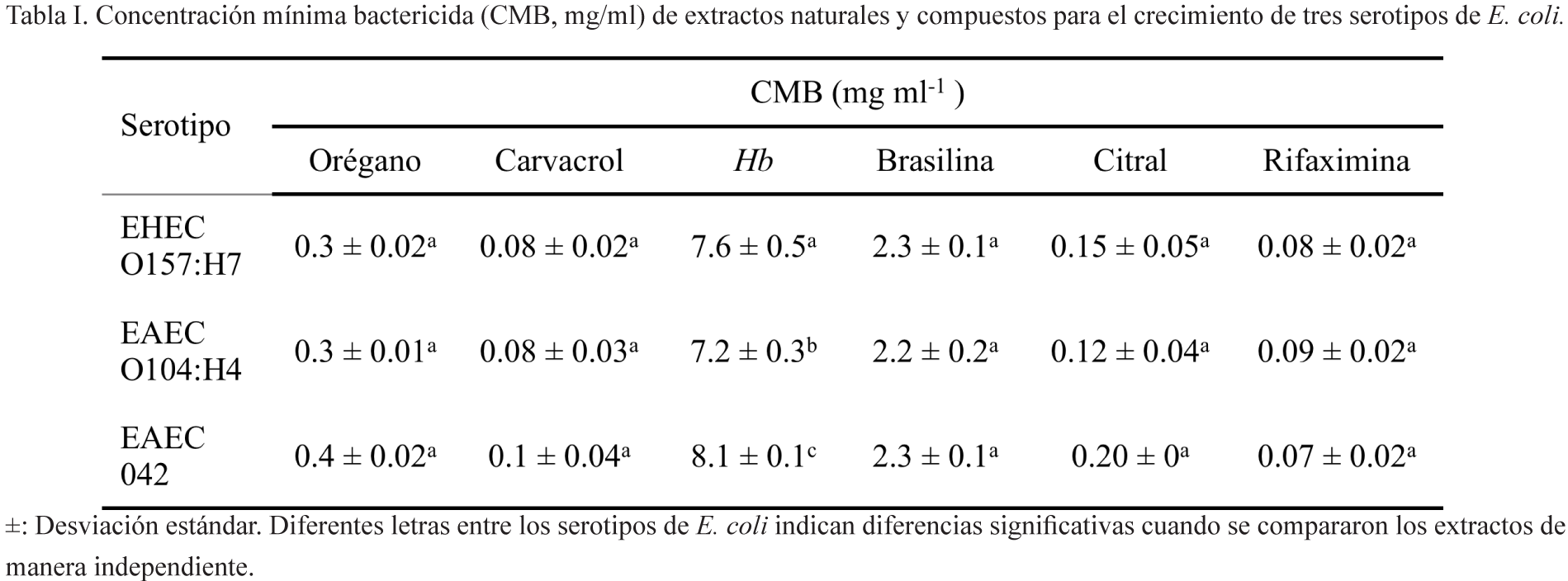
Determinación de la concentración mínima bactericida (CMB)
La CMB se determinó por el método de microdilución (García-Heredia et al., 2016). Brevemente, pocillos de microplacas estériles de 96 pozos se llenaron con 100 μl de caldo MH 2X adicionando 100 μl de diferentes concentraciones de los compuestos a probar. Cada pocillo fue inoculado con 20 μl de la suspensión bacteriana (2 x 106 UFC) y se incubaron a 37°C/24 h. Una alícuota de cada pocillo se sembró por estría sobre placas con agar MH, y se incubaron a 37°C/24 h. La CMB se estableció como la concentración más baja que no permitió crecimiento visible en las placas de agar MH.
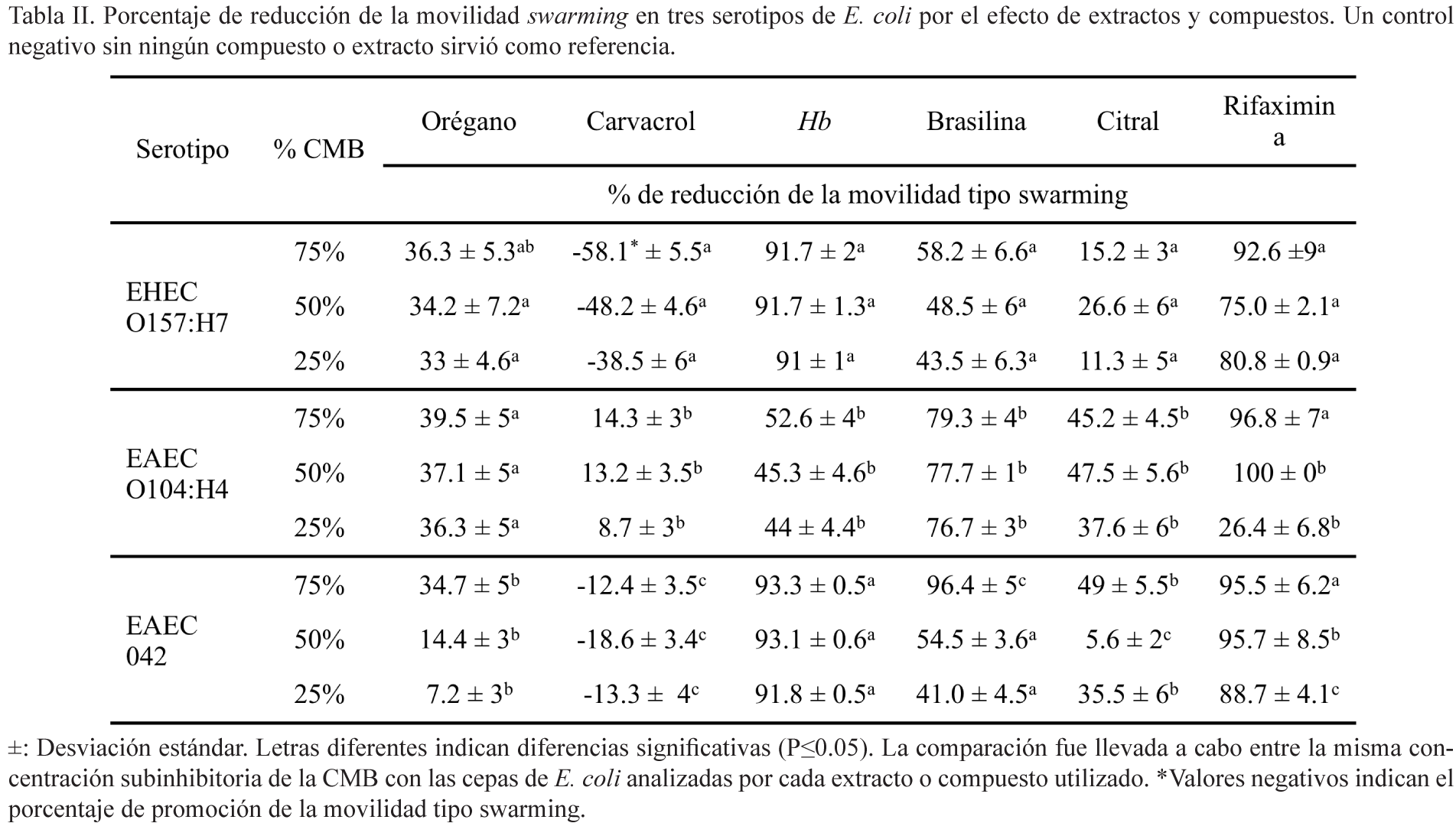
Efecto de los compuestos o extractos sobre la movilidad tipo swarming
La movilidad tipo swarming se determinó por el método descrito por Lai et al. (2009) con algunas modificaciones. Brevemente, 5 μl de los cultivos bacterianos (1 x 108 CFU/ml) se inocularon en el centro cajas Petri con agar LB suave (0.35% agar) adicionado con concentraciones de los compuestos a probar y se incubaron a 37°/24 h. La magnitud de la movilidad tipo swarming se basó en la distancia de migración a partir del centro de la placa (García-Heredia et al., 2016).
Formación de biopelícula
La formación de biopelícula fue estudiado por el método de Yadav et al. (2015). Placas de microtitulación estériles se llenaron con 200 μl de caldo LB con glucosa a 0.2%, citrato de sodio a 1% (promotores de la formación de biopelícula) y los compuestos a analizar. Cada pocillo se inoculó con 20 μl de la cepa activada (1 x 106 UFC) y la placa se incubó a 37°C/24 h. Después de medir la absorbancia (630 nm) de cada pocillo, los sobrenadantes se eliminaron y los pocillos se lavaron con agua destilada estéril y se secaron al aire. Las bacterias que permanecieron en los pocillos se tiñeron con 200 μl de cristal violeta a 0.1% (CV) e incubaron a 37°C/15 min. Después, las placas se lavaron con agua destilada para finalmente añadir 200 μl de etanol a 96% a fin de disolver el CV dentro de la matriz de la biopelícula. Después de 5 min de incubación a 37°C, se midió la absorbancia (570 nm) a cada pocillo.
El índice de formación de biopelícula (BFI por sus siglas en inglés) se calculó con la siguiente fórmula:
BFI = (AB – CW)/G
donde AB corresponde a la absorbancia de las bacterias pegadas teñidas, CW es la absorbancia de los controles teñidos (con los compuestos a analizar) y G es la absorbancia de la bacteria que se encontraba en suspensión. Los valores de BFI >1.10 se interpretaron como una biopelícula fuerte, de 0.70 a 1.09 como una biopelícula moderada, de 0.35 a 0.69 como una biopelícula débil y <0.35 no formación de biopelícula (García-Heredia et al., 2013).
Análisis de la expresión génica
Las bacterias se cultivaron toda la noche a 37°C y una alícuota (400 μl) se inoculó en tubos con 4 ml de caldo LB conteniendo los compuestos a probar a varias concentraciones. Los tubos se incubaron a 37°C/3 h en agitación. Inmediatamente después el ARN se extrajo y purificó usando el sistema Isolate II RNA Mini Kit y la integridad del ARN obtenido se determinó mediante un espectrofotómetro UV-Vis. El ADNc se sintetizó usando el sistema iScriptTM cDNA Synthesis. Finalmente, se llevó a cabo la reacción de PCR de tiempo real utilizando el sistema iQ SYBR Green Supermix. Los iniciadores utilizados incluyeron, para el gen pic (mediador de colonización, presente en O104:H4 y 042; Safadi, et al., 2012), aggR (regulador maestro del plásmido de adherencia agregativa, presente en O104:H4 y 042; Safadi, et al., 2012), rpoS (relacionado al estrés, presente en las tres cepas analizadas; Liu, et al., 2010), Stx2 (Sigatoxina, presente en O157:H7 y O104:H4; Safadi, et al., 2012). El gliceraldehydo-3-fosfato deshidrogenasa (GAPDH), un gen constitutivo, fue utilizado como normalizador y las bacterias sin tratamientos se usaron como controles y como niveles basales para comparar la expresión génica.
Análisis estadístico
Todos los ensayos se realizaron al menos dos veces cada uno por duplicado o triplicado. La comparación entre tratamientos se analizó por la prueba Fisher LSD usando el paquete computacional GraphPad Prism versión 6.c for IOX. El nivel de significancia utilizado fue de 95%.
RESULTADOS
CMB de los extractos y compuestos
Todos los extractos y compuestos analizados mostraron actividad antimicrobiana contra todas las cepas analizadas (tabla I). El carvacrol mostró la mayor actividad antimicrobiana, resultando valores de CMB similares a los mostrados por la rifaximina (control positivo), seguido de orégano, brasilina y Hb. No se encontraron diferencias significativas (P≤0.05) en las CMB de cada extracto o compuesto (orégano, carvacrol, brasilina, citral y rifaximina) entre las cepas analizadas.
Efecto en la movilidad tipo swarming
Se observó mucha variabilidad en la movilidad tipo swarming por las tres cepas analizadas, cuando se pusieron en contacto con los diferentes extractos y compuestos. De manera muy interesante, la cepa EAEC O104:H4 mostró un comportamiento más parecido al mostrado por la EHEC O157:H7 en algunos tratamientos, en otros fue semejante al de la cepa EAEC 042, y en ocasiones totalmente diferentes a las anteriores (tabla II). El orégano a sus diferentes concentraciones redujo la movilidad tipo swarming de 7 a 39% en las cepas analizadas. Sin embargo, de manera sorpresiva, el carvacrol promovió la movilidad en EHEC O157:H7 y EAEC 042, pero la redujo, aunque de manera muy leve, en la cepa EAEC O104:H4. En el caso de los extractos de Hb, las cepas EHEC O157:H7 y EAEC 042 disminuyeron el swarming en niveles arriba de 90%, en tanto que de entre 40 y 50% para la cepa EAEC O104:H4. Por su parte, la brasilina redujo este tipo de movilidad en 49% para EHEC O157:H7, 63% en EAEC 042 y 77% en EAEC O104:H4. Además, el citral también provocó reducción de la movilidad de 5 a 49% dependiendo de la cepa, mientras que la mayoría de las concentraciones de rifaximina utilizadas provocaron reducciones que fueron entre 75 a 100%, con excepción de cuando se agregó 25% de la CMB en EAEC O104:H4, que sólo mostró una reducción de la movilidad de 26%.
Formación de biopelícula
Como en el caso de la movilidad swarming, se observó alta variabilidad en el BFI cuando las células se trataron con los extractos o compuestos (tabla III). Se observó disminución del BFI (de 32 a 41%) en EAEC 042 por orégano y carvacrol a 75% de la CMB. Hb provocó una disminución (29 a 82%, P≤0.05) del BFI en todas las cepas estudiadas, y la brasilina produjo una disminución del BFI (58 y 64%) en EHEC O157:H7 y EAEC 042, respectivamente. Citral redujo (40%) el BFI en EHEC O157:H7 cuando se le agregó 75% de la CMB y en EAEC 042 en todas las concentraciones probadas. De manera general, la formación de biopelícula por EAEC 042 se redujo al agregar la mayoría de los extractos/compuestos analizados. Es importante mencionar que al aplicar 25% de sus CMBs, el orégano y el carvacrol incrementaron casi al doble el BFI de E. coli O157:H7, y que la brasilina y el citral incrementaron el BFI (46 y 53%, respectivamente) en EAEC O104:H4. Además, la rifaximina indujo el BFI cuando se agregó a concentraciones correspondientes a 50 y 75% de la CMB de EHEC O157:H7 y a 75% de la CMB en EAEC O104:H4.

Expresión de genes
Se analizó la influencia de los extractos o compuestos a una concentración equivalente a 75% de la CMB en la expresión de tres genes de virulencia y el factor rpoS en las cepas, encontrándose que el efecto fue cepa y com- puesto dependiente y no siguieron un patrón consistente en las cepas EHEC O157:H7 y EAEC O104:H4. Solamente en el caso de la cepa EAEC042 se observo un patrón constante en el que se disminuía la expresión de los genes pic y rpoS, y un incremento del regulador maestro aggR (tabla IV) en la mayoría de los tratamientos al comparase con el control. La expresión del gen Stx2 en EHEC O157:H7 y EAEC O104:H4, disminuyó de 34% a 96%, al exponerse a los compuestos o extractos analizados. Sin embargo, la brasilina incrementó su expresión (entre 30 y 40%) en las dos cepas, en tanto que el carvacrol sólo lo hizo en la cepa EAEC O104:H4.
La mayoría de los extractos incrementaron la expresión del regulador maestro aggR en EAEC O104:H4, con excepción de orégano y citral que redujeron su expresión entre 40 y 60%. En el caso de la cepa EAEC 042, este gen se sobreexpresó de 10 a 550% con los extractos o compuestos agregados cuando se comparó con el control. El gen pic, presente en las cepas EAEC, se sobreexpresó entre 20 y 110% con todos los extractos o compuestos agregados en EAEC O104:H4, con excepción de orégano y citral que incrementaron la expresión del gen entre 30 y 60%. Por el otro lado, la expresión del gen pic se redujo entre 50 a 97% al agregar los tratamientos a la cepa EAEC 042.
La expresión de rpoS se incrementó en EHEC O157:H7 con orégano, carvacrol y rifaximina, pero se redujo por Hb y citral. La brasilina incrementó la expresión de este gen en EAEC O104:H4. De manera interesante, todos los extractos y compuestos redujeron la expresión (20 a 60%) de EAEC 042 cuando se comparó con el control (tabla IV).
DISCUSIÓN
Numerosas plantas comestibles y sus compuestos purificados han mostrado tener actividad antimicrobiana que puede deberse a mecanismos diversos, como la ruptura de la membrana celular, incremento en la permeabilidad membranal, disminución del pH intracelular, hiperpolarizaión de la membrana celular, disminución del nivel intracelular de ATP, etcétera (Sánchez et al., 2010). Se ha determinado que el carvacrol tiene la capacidad de unirse al ADN (Nafisi et al., 2004), sugiriendo que la formación de complejos de ADN puede ser un mecanismo para las interacciones antimicrobiano-ADN.
En este estudio, los compuestos aislados carvacrol y brasilina demostraron tener una mayor actividad bactericida en comparación con los extractos completos de orégano y Hb, respectivamente. Estas observaciones son consistentes con estudios previos que han señalado que el carvacrol presenta mayor efecto bactericida cuando se compara con extractos de orégano. Se piensa que la actividad antimicrobiana es debida a la interacción del compuesto con la bicapa lipídica de la membrana celular, provocando pérdida de la integridad y como consecuencia fuga del material celular (Nostro et al., 2007).
Se ha reportado actividad antimicrobiana del citral contra E. coli (Somolinos et al., 2009), y en este trabajo también se encontró que posee actividad contra cepas patógenas de E. coli. En este estudio también confirmamos la actividad antimicrobiana de la rifaximina contra E. coli, tal como había sido reportado previamente (Hopkins et al., 2014).
En este estudio, casi todos los tratamientos redujeron la movilidad tipo swarming en las cepas de E. coli analizadas, sin embargo, contrario a nuestras expectativas, el carvacrol la promovió en EHEC O157:H7 y EAEC 042, pero no en EAEC 0104:H4. Entre las posibles causas de estos resultados están variaciones fisiológicas y morfológicas de los aislados y a la vez la acción de los diferentes antimicrobianos naturales. Estudios previos han señalado que el carvacrol puede reducir la movilidad de Salmonella Typhimurium (Van Alphen et al., 2012). Además, está establecido que las células nadadoras tienen gran actividad metabólica y, como consecuencia, pudieran desplegar un fenotipo resistente contra antimicrobianos (Lai et al., 2009). Más aún, se ha establecido que B. cereus tiene la capacidad de adaptarse a concentraciones subletales de carvacrol debido a variaciones en sus ácidos grasos, disminuyendo la fluidez de la membrana (Ultee et al., 2000). Se ha reportado que el citral es capaz de causar daños subletales a células de E. coli (Somolinos et al., 2009); sin embargo, las células se pueden recobrar mediante la síntesis de lípidos, requiriendo para esto grandes cantidades de energía que, en un momento dado, pudieran afectar el desarrollo de células híper flageladas, y como consecuencia, reducir la movilidad tipo swarming.
El efecto observado de los compuestos utilizados en la producción de biopelícula varió grandemente entre las cepas y los compuestos probados. Villegas et al. (2015) han reportado que la rifaximina a concentraciones de 0.032mg/ml y 32 mg/ml disminuyeron la formación de biopelícula en algunas cepas STEC. En nuestro estudio, la concentración más baja de rifaximina que utilizamos (25% de la CMB) fue capaz de provocar una reducción de la biopelícula en la cepa EAEC042. Sin embargo, a 75% de la CMB, contrario a lo esperado, promovió su desarrollo en EHEC O157:H7 y EAEC O104:H4, dando muestras de las diferencias entre serotipos.
A bajas concentraciones (25% de la CMB) de orégano, carvacrol, brasilina y citral se produjo un incremento en la formación de biopelícula en EHEC O157:H7 y EAEC O104:H4. Previamente se ha reportado que a concentraciones subletales de antibióticos se puede inducir la formación de biopelícula en algunas bacterias. Esto se debe a una respuesta de protección que inducen alteraciones a nivel del monofosfato cíclico de diguanosina (Hoffman et al., 2005).
Es de suma importancia indicar que la mayoría de los compuestos analizados en este estudio disminuyeron la expresión del gen Stx2 en EHEC O157:H7 y EAEC O104:H4. Estos compuestos se suman a la rifaximina que disminuye la inducción de Stx2 y ha sido usada para el tratamiento de brotes de enfermedad (Bielaszewska et al. 2012).
Los productos codificados por el gen rpoS pueden funcionar como represores de la movilidad tipo swarming en Serratia sp. (Wilf y Salmond, 2012), por lo que en este trabajo examinamos si existía una relación entre la expresión de rpoS, movilidad tipo swarming y formación de biopelícula por efecto de los compuestos utilizados. Para el caso de EHECO157:H7, la expresión de rpoS se incrementó por la mayoría de los compuestos analizados, con excepción de Hb y citral, sin embargo, la movilidad se redujo con la mayoría de los compuestos, menos con carvacrol; por lo que el incremento en la expresión de rpoS no se correlacionó con la disminución de swarming. De manera similar, la formación de biopelícula se redujo con Hb, brasilina y citral, pero incrementó con rifaximina, por lo que no existió una relación entre los compuestos analizados, la expresión de rpoS y la formación de biopelícula.
Cuando estudiamos la cepa EAEC O104:H4, la expresión de rpoS se incrementó por orégano, carvacrol y rifaximina, pero se redujo al añadir Hb y brasilina. Más aún, la movilidad tipo swarming se redujo en todos los tratamientos, pero la producción de biopelícula sólo se disminuyó con Hb y se incrementó con rifaximina. La expresión de rpoS se redujo con todos los tratamientos en EAEC 042, en tanto que la biopelícula se redujo o no se afectó y la movilidad tipo swarming disminuyó con todos los tratamientos menos carvacrol. Con base en lo anterior, no existió una relación clara entre la expresión del gen rpoS y los parámetros analizados en este estudio en las cepas EAEC analizadas.
Una observación interesante en este estudio fue que en EAEC O104:H4 los niveles de expresión de pic y rpoS fueron variables, mientras que en la cepa EAEC 042, la expresión de ambos genes fue suprimida al agregar los compuestos o extractos analizados. Esta observación indica que, aunque ambas son cepas pertenecientes al patotipo enteroagregativo (EAEC), existen diferencias en su regulación génica, sugiriendo que la respuesta mostrada a los antimicrobianos, además de ser dependientes del compuesto, lo es de las variaciones entre las cepas. Adicionalmente, se demostró que existen grandes variaciones en la expresión de genes cuando se utilizaron extractos crudos y sus compuestos mayoritarios. Estas diferencias pueden deberse a la presencia de compuestos no identificados en los extractos crudos que pueden modificar su actividad (Ávila-Sosa et al., 2010).
*Universidad Autónoma de Nuevo León
Contacto: norma@microbiosymas.com
REFERENCIAS
Ávila-Sosa, R., et al. (2010). Extracts of Mexican oregano (Lippia berlandieri Schauer) with antioxidant and antimicrobial activity. Food Bioprocess Technol. 3, 434-440.
Bielaszewska, M., et al. (2012). Effects of antibiotics on Shiga toxin 2 production and bacteriophage induction by epidemic Escherichia coli O104:H4 strain. Antimicrob. Ag. Chemother. 56, 3277-3282.
Boisen, N., et al. (2014). The Presence of the pAA plasmid in the German O104:H4 Shiga toxin type 2a (Stx2a)-producing Enteroaggregative Escherichia coli strain promotes the translocation of Stx2a across an epithelial cell monolayer. J. Infect. Dis. 210, 1909-1919.
Fouladkhah, A., et al. (2013). Biofilm formation of O157 and non-O157 shiga toxin-producing Escherichia coli and multidrug-resistant and susceptible Salmonella Typhimurium and Newport and their inactivation by sa- nitizers. J. Food Sci. 78, 880-886.
García-Heredia, A., et al. (2013). Efficacy of citrus-based disinfectants to inhibit growth, swarming, and biofilm formation of Salmonella and decontaminate pars- ley. J. Food Agri. Environ. 11, 1295-1299.
García-Heredia, A., et al. (2016). Natural plant products inhibits growth and alters the swnming motility, blu film formation, unexpression of virulence genes in enteroaggregative and entero hemorrhagic Escherichia coli. Food Microbiology. 59:124-132
Guy, L., et al. (2013). Adaptive mutations and replacements of virulence traits in the Escherichia coli O104:H4 outbreak population. PLoS ONE. 8, e63027.
Heredia, N., et al. (2005). Extracts of Haematoxylon brasiletto inhibit growth, verotoxin production, and adhesion of enterohemorrhagic Escherichia coli O157:H7 to HeLa cells. J. Food Prot., 68, 1346-1351.
Hoffman, L.R., et al. (2005). Aminoglycoside antibiotics induce bacterial biofilm formation. Nature. 436, 1171-1175.
Hopkins, K.L., et al. (2014). In vitro activity of rifaximin against clinical isolates of Escherichia coli and other enteropathogenic bacteria isolated from travelers returning to the UK. Int. J. Antimicrob. Ag. 43, 431- 437.
Kim, S.Y., et al. (2011). Antimicrobial activity of plant extracts against Salmonella Typhimurium, Escherichia coli O157:H7, and Listeria monocytogenes on fresh lettuce. J. Food Science. 76, 41-46.
Lai, S., et al. (2009). Swarming motility: a multicellular behaviour conferring antimicrobial resistance. Environm. Microbiol. 11, 126-136.
Law, D. (2000). Virulence factors of Escherichia coli O157:H7 and other Shiga toxin-producing E. coli. J. App. Microbiol. 88, 729-745.
Liu, Y., et al. (2010). Quantification of viable but nonculturable Escherichia coli O157:H7 by targeting the rpoS. Anal. Chem. 82, 2612-2615.
Melton-Celsa, A.R. (2014). Shiga toxin (Stx) classification, structure, and function. Microbiol. Spectrum. 2, 37-53.
Nafisi, S., et al. (2004). Thymol and carvacrol binding to DNA: model for drug-DNA interaction. Biopolymers. 74, 345-351.
Nostro, A., et al. (2007). Effects of oregano, carvacrol and thymol on Staphylococcus aureus and Staphylococcus epidermidis biofilms. J. Med. Microbiol. 56, 519-523.
Orue, N., et al. (2013). Decontamination of Salmonella, Shigella and Escherichia coli O157:H7 from leafy green vegetables using edible plant extracts. J. Food Sci. 78, 290-296.
Quintanilla-Licea, R., et al. (2014). Antiprotozoal Activity against Entamoeba histolytica of plants used in Northeast Mexican traditional medicine. Bioactive compounds from Lippia graveolens and Ruta chalepensis. Molecules. 19, 21044-21065.
Rivero-Cruz, J.F. (2008). Antimicrobial compounds isolated from Haematoxylon brasiletto. J. Ethnopharmacol. 119, 99–103.
Sánchez, E., et al. (2010). Extracts of edible and medicinal plants damage membranes of Vibrio cholerae. Appl. Environ. Microbiol. 76, 6888-6894.
Somolinos, M., et al. (2009). Inactivation of Escherichia coli by citral. J. Appl. Microbiol. 108, 1928-1939.
Ultee, A., et al. (2000). Adaptations of the food-borne pathogen Bacillus cereus to carvacrol. Arch Microbiol. 174, 233-238.
Van Alphen, et al. (2012). The natural antimicrobial carvacrol inhibits Campylobacter jejuni motility and infection of epithelial cells. PLoS ONE, 7, e45343.
Villegas, N.A., et al. (2015). Effect of antibiotics on cellular stress generated in Shiga toxin-producing Escherichia coli O157:H7 and non-O157 biofilms. Toxicol. in Vitro. 29, 1692-1700.
Wilf, N.M., y Salmond, G.P.C. (2012). The stationary phase sigma factor, RpoS, regulates the production of a carbapenem antibiotic, a bioactive prodigiosin and virulence in the enterobacterial pathogen Serratia sp. ATCC 39006. Microbiol. 158, 648-658.
Yadav, M.K., et al. (2015). Eugenol: A phyto-compound effective against methicillin-resistant and methicillin-sensitive Staphylococcus aureus clinical strain biofilms. PLoS ONE, 10, e0119564.
Recibido: 8 de septiembre de 2017
Aceptado: 13 de octubre de 2017