El golfo de California: un océano joven, región megadiversa, vínculo entre tectónica y ecología

Thierry Calmus*, Alberto Búrquez**, Angela Martínez Yrízar**
CIENCIA UANL / AÑO 20, No. 85, julio-septiembre 2017
Para los geólogos, los océanos se distinguen de los continentes por la naturaleza de la corteza que constituye su basamento. La corteza oceánica es más densa que su equivalente continental, y por lo tanto se encuentra más hundida en el manto subyacente que la sostiene. La corteza continental, la oceánica y el manto superior rígido forman la litósfera. La profundidad de equilibrio cuando la litósfera oceánica está fría es de aproximadamente 4,500 m. Siendo más densas, las regiones de la Tierra con una litósfera oceánica forman cuencas que contienen 97% del volumen de agua disponible en la superficie de la Tierra. Así, la distribución geográfica del agua de mar depende de la repartición de litósfera oceánica. Por su densidad, la litósfera oceánica tiene la posibilidad de hundirse en el manto inferior y desaparecer paulatinamente por el fenómeno de subducción. Ya que el volumen y la superficie de la Tierra son constantes, la subducción se acompaña de creación de nueva litósfera oceánica a lo largo de las dorsales oceánicas a partir de material magmático que proviene de la astenósfera, una de las capas del interior de la Tierra. Estos fenómenos tectónicos han sido cuantificados desde hace 50 años en el marco de la tectónica de placas, la cual unifica los diferentes fenómenos geológicos (magmáticos, volcanológicos, sismológicos, orogénicos, etcétera) en un sistema único, global. Este paradigma se basa en el hecho de que la Tierra es un planeta activo en el cual existen intercambios de calor entre sus diferentes capas desde el núcleo interno hasta la superficie. En particular, el manto inferior que constituye la parte mediana de la Tierra tiene un comportamiento plástico, lo cual permite un intercambio de calor por convexión, mientras que la parte superficial, que es rígida, está dividida en varias placas litosféricas que se mueven horizontalmente, unas con respecto a otras, en un movimiento continuo.
Para los biólogos, la separación entre océanos y continentes puede ser tan simple como la presencia o ausencia de grandes masas de agua superficial, o tan compleja como distinguir entre los componentes biológicos que integran los diversos ecosistemas marinos y terrestres; sistemas dinámicos que presentan interacciones a diferentes escalas temporales y espaciales. Los fenómenos geológicos afectan de manera decisiva los componentes biológicos, tanto aquellos de carácter oceánico como los de la contraparte terrestre; es decir, existe un estrecho vínculo entre la tectónica y la diná- mica ecológica. En las dorsales del océano profundo prosperan ecosistemas únicos que dependen de la síntesis de materia orgánica a partir de la energía de enlaces químicos presentes en los minerales que emiten las ventilas hidrotermales. Estas ventilas o fumarolas submarinas forman chimeneas compuestas principalmente de anhidrita, un mineral compuesto de sulfato de calcio anhidro. Fueron descubiertas en 1977 por investigadores de la Institución Oceanográfica de Woods Hole y están presentes en las dorsales oceánicas, incluyendo pequeños segmentos de la dorsal del Golfo de California. En estas chimeneas, los sulfuros de cobre, hierro y zinc se precipitan y son la base del metabolismo de las arqueas, un tipo muy especial de bacterias quimiosintéticas que soportan una cadena alimenticia que incluye organismos muy especializados como los gusanos gigantes de tubo, diversas especies de almejas, cangrejos y camarones adaptados a la vida en estos oasis submarinos profundos.
En las zonas abisales existe también una variada fauna que depende de la “lluvia” de materia orgánica –restos de plancton, peces y sedimentos continentales– que cae continuamente desde las aguas superficiales e intermedias al fondo del mar. En cambio, en las aguas más someras dentro de la zona fótica, que varía desde una delgada lámina superficial hasta profundidades de cerca de 200 m o hasta donde penetra la luz del sol, prosperan los ecosistemas más diversos y productivos del mundo marino. Los sargazos en los mares cálidos y las macroalgas (kelp) en los más fríos forman verdaderos bosques con muy alta biodiversidad y productividad biológica. En las zonas litorales plantas como Zoostera y Thalassia forman extensos pastizales marinos.
La interfase entre el mar y la tierra forma una angosta faja donde se desarrollan ecosistemas híbridos altamente dinámicos dominados por procesos geomorfológicos donde prevalece por un lado la acción de las mareas y el oleaje y por otro los aportes de agua dulce y sedimentos continentales. Destacan los manglares, los sistemas deltaicos y estuarinos con extensos sistemas de marismas, las salinas con una flora y fauna sumamente especializada, las playas arenosas asociadas en mayor o menor grado a sistemas de dunas, y en las zonas desérticas, los estuarios hipersalinos con condiciones de elevada salinidad y escaso aporte de agua dulce. Estos ambientes litorales representan los lugares de anidación y crecimiento de numerosas especies marinas, incluyendo peces, moluscos, crustáceos, anélidos e incontables aves playeras, residentes y migratorias (Carmona et al., 2015).
Entorno geográfico
El Golfo de California, también llamado Mar de Cortés (originalmente llamado Mar Bermejo por la coloración de sus aguas durante las avenidas del río Colorado y después bautizado Mar de Cortez por Francisco de Ulloa en 1539) es un largo y angosto mar interior subtropical de más de 1,200 km de longitud, una anchura entre 80 y 200 km y una superficie aproximada de 160,000 km2 . La parte sur del golfo presenta relieves submarinos abruptos con fosas abisales de más de 3 km de profundidad, mientras que su parte septentrional es somera, lo que ocasiona una oscilación de mareas que llegan a superar los 9 m. El golfo incluye casi 1,000 islas entre las que destacan las dos islas más grandes de México: Ángel de la Guarda y Tiburón. Por su alta productividad biológica, sus aguas profundas, arrecifes rocosos y playas arenosas, Jacques-Yves Cousteau llamó al Golfo de California “el acuario del mundo”. Está rodeado por los estados de Baja California, Baja California Sur, Sonora, Sinaloa y Nayarit. Sus costas albergan casi 8 millones de personas entre las que se incluyen los grupos indígenas, los Tohono O’odham, Comcaac, Cucapá, Yoreme, Yoeme, Kikapu, Kiliwa y Cora (Luque et al., 2016), además de los grupos ya extintos Cochimí y Pericú de la península. Actualmente, importantes poblaciones bordean sus costas y el Golfo de California representa la región más productiva del país desde el punto de vista pesquero (Arreguín-Sánchez y Arcos-Huitrón, 2011).
Evolución geológica
Hace 12 Ma, la subducción de la placa Farallón, debajo de la placa Norteamericana y la colisión de la dorsal o su acercamiento con la placa Norteamericana, provocó un cambio cinemático y el inicio de un desplazamiento lateral derecho de la placa Pacífico con respecto a la placa Norteamericana. En otras palabras, a partir de esa fecha, la placa Pacífico empezó a moverse hacia el nornoroeste con respecto a su placa vecina. Al principio, el desplazamiento se ejercía a lo largo del antiguo límite de la placa Farallón y de la placa Norteamericana, pero unos millones de años después, este límite cruzó hacia dentro del continente, en la región que corresponde a lo que paulatinamente se transformaría en el Golfo de California. La parte continental correspondiente a la futura península de Baja California se incorporó a la placa Pacífico y comenzó a migrar de forma solidaria hacia el nornoroeste junto con la placa Pacífico. El desprendimiento de la península tuvo por consecuencia un estiramiento y un adelgazamiento progresivo de la litósfera continental preexistente que permitió, hace 3.6 Ma (DeMets, 1995), la formación de nueva corteza oceánica localizada en pequeñas cuencas en la parte sur del golfo (cuencas de tipo “pull apart”), como las cuencas de Alarcón y la de Guaymas (figura 1).
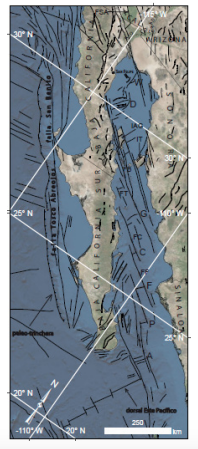
Figura 1. Marco tectónico regional del Golfo de California
en el cual se observan las principales estructuras tectónicas de la margen oeste de la península de Baja California (Paleo-trinchera
y sistema de fallas Tosco-Abreojos-San Benito), y del propio Golfo (fallas transformantes y cuencas de tipo “pull apart”). CAL: Estado de California.
Fallas transformantes:
FSA: Falla San Andrés; FB:
Falla Ballenas; FT: Falla Tortuga; FC: Falla Carmen; FF: Falla Farallón; Cuencas de tipo “pull apart”: W: cuenca Wagner; D: cuenca Delfín; G: cuenca Guaymas; C: cuenca Carmen; F: cuenca Farallón; P: cuenca Pescadero; A: cuenca Alarcón.
Si observamos más a detalle, este movimiento presenta una ligera componente de extensión (Antonelis et al., 1999), que lentamente separa la península de Baja California del continente. Sin embargo, la componente lateral ha prevalecido durante los últimos 6 millones de años, y el desplazamiento acumulado corresponde a una distancia de 240 km (Oskin et al., 2001; Oskin y Scott, 2003). La frontera entre las placas Pacífico y Norteamericana está conformada por una sucesión de fallas laterales en escalón desde el desemboque del Golfo de California hasta la falla San Andrés en California. En el relevo entre dos fallas, se desarrollan las cuencas que mencionamos anteriormente (figura 1).
Como en cualquier límite de placas, las fallas del Golfo de California generan sismos que corresponden a una relajación elástica de la corteza terrestre sometida a esfuerzos tectónicos. La distribución de los hipocentros (lugar de ruptura) de los sismos, así como los mapas batimétricos permiten trazar la trayectoria de las fallas. El estudio de los sismos y las mediciones GPS permiten calcular una velocidad promedio de 5 cm por año (Dixon et al., 2000); una distancia de 24 m, desde que en 1539 Francisco de Ulloa, en la búsqueda del estrecho de Anián, dio su nombre a este mar que separaba Nueva Galicia de lo que se llamaría después California.
Este cálculo realizado durante las dos últimas décadas con instrumentos GPS confirma las evaluaciones realizadas a partir de diversas evidencias geológicas; por ejemplo, la correlación entre rocas volcánicas de 12 Ma que se encuentran en el Cerro Eucla al norte de Bahía Kino, en la Sierra Kunkaak en la Isla Tiburón y en la región de San Felipe en Baja California (Oskin et al., 2001). La edad de las rocas volcánicas contemporáneas, de la ruptura continental y de los primeros sedimentos, permite también precisar la evolución geológica de las primeras cuencas a lo largo de la costa este de la península (Ortlieb y Colletta, 1984; Holt et al., 2000).
La ruptura continental modificó la geografía regional permitiendo la incursión progresiva de sur a norte de aguas marinas del golfo en formación. Nace un nuevo mar, somero al inicio, que se profundiza paulatinamente conforme aumenta la separación y la subsidencia asociada a la extensión de la corteza (Francheteau et al., 1984). A la par, la fisionomía de la red fluvial de la Sierra Madre Occidental cambia de dirección; los principales drenajes que originalmente fluían hacia la región del sur de Sinaloa, Nayarit o Jalisco encuentran escape hacia el protogolfo. Los sedimentos marinos fosilíferos que se encuentran a lo largo de las costas del Golfo (Helenes y Carreño, 1999; Ochoa-Landín et al., 2000) apoyan la hipótesis de una transgresión marina desde el sur.
Evolución biológica
La génesis del Golfo de California llevó a la creación de ecosistemas marinos y terrestres únicos. Por un lado, la creación de un mar interior muy cálido (figura 2) con fuertes oscilaciones en la salinidad y enormes aportes de sedimentos en su extremo septentrional y por otro, el aislamiento de los componentes biológicos que emigraron junto con la península separándose de sus contrapartes continentales. En el mar profundo, la apertura de ventilas hidrotermales asociadas a la creación de piso oceánico, en las cuencas “pull apart” del sur del Golfo, originó oasis muy diversos. La migración de la península no sólo permitió la creación de nuevo fondo marino, sino que protegió al nuevo mar interior de la corriente fría de California que desciende desde el estrecho de Bering. Asimismo, las variaciones bruscas de profundidad, la ubicación de grandes islas en la cintura del Golfo (“Midriff Islands”) y las diferencias de temperatura y salinidad, permiten un sistema de corrientes complejo que provoca surgencias que acarrean nutrientes desde las profundidades y enriquecen las aguas superficiales, permitiendo una enorme productividad biológica sólo equiparable por otro mar con características muy similares: el Mar Rojo.
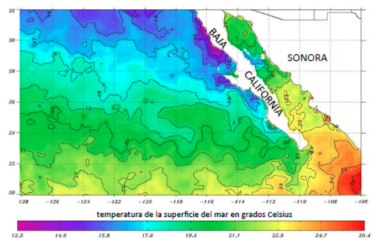
Figura 2. Distribución de la temperatura superficial del mar entre las latitudes 20 y 32ºN y las longitudes 106 y 128ºW, el 15 de abril de 2017. La temperatura es una mezcla que combina datos provenientes de todos los satélites geoestacionarios y de orbita polar, de Estados Unidos, de Japón y de Europa. Para generar un producto diario con
una resolución espacial de 5 km, se usa la resolución espacial más alta de los satélites de orbita polar que es de 1 km con observaciones cada seis horas aproximadamente, combinada con la resolución temporal más alta de los satélites geoestacionarios de aproximadamente
5.5 km con observaciones cada hora. Se observa el fuerte contraste de temperaturas entre el Golfo de California y el Pacífico. Por ejemplo, la isoterma 23ºC en el Golfo se encuentra aproximadamente 600 km más al norte que en el Pacífico. Modificado a partir de NOAA/NESDIS (National Orceanic and Atmospheric Organization/National
Environmental Satellite, Data and Information Service).
Desde su apertura hace 5-6 Ma, el Golfo es una de las regiones marinas más ricas del mundo (Felger et al., 2013). Se calcula que contiene más de 8,000 especies animales. Existen más de 4,900 especies de invertebrados, aunque de acuerdo a Brusca y Hendrickx (2010) la diversidad total puede exceder las 7,000 especies. De las casi 1,000 especies de peces, 87% está dominado por especies de aguas tropicales y 10% son endémicas, esto es, sólo se encuentran aquí (Hastings et al., 2010). En el Golfo existen cinco de las siete especies de tortugas marinas del mundo, todas ellas consideradas como amenazadas o en peligro (Márquez, 1990; Seminoff, 2010). Los mamíferos marinos son también sorprendentemente diversos ya que se tienen registradas 36 especies, de las cuales 31 son cetáceos (39% de los cetáceos del mundo). Entre éstas, la vaquita (Phocaena sinus) está a punto de extinguirse y cuatro especies están catalogadas como en peligro: las ballenas azul, sei, de aleta y franca. Además, existen cuatro especies de pinnípedos (focas, elefantes marinos y dos especies de lobo marino; Urban, 2010). De aves marinas, 56 especies habitan en el Golfo de California; seis de ellas son cuasiendémicas, diez son residentes (pasan todo su ciclo de vida en el golfo), 26 son migratorias (usan el golfo como parte del corredor migratorio) y 18 usan las islas como sitios de reproducción (Velarde y Anderson, 1993).
En la porción terrestre, especialmente en la peninsular, existen numerosas especies endémicas, tanto de plantas como de animales (figura 3). Estos endemismos son notables, no sólo por el efecto del aislamiento peninsular que cortó el flujo genético con las poblaciones continentales debido a la migración de la península, sino también porque su estudio permite la inferencia de procesos geológicos específicos que dejaron una marca biológica. Para citar sólo un ejemplo, el trabajo de Riddle et al. (2000) permitió inferir la historia geológica de la península desde el Mioceno Tardío hasta el Pleistoceno a través del estudio de 12 grupos de especies de mamíferos, aves, reptiles y anfibios. Estos autores encontraron que la arquitectura filogeográfica (esto es, las relaciones de parentesco entre especies en el entorno geográfico) sólo puede explicarse por una serie de eventos vicariantes (es decir, de diferenciación entre linajes por el aislamiento geográfico) debidos a: 1) un canal marino transpeninsular durante el Pleistoceno Medio, 2) una transgresión de la porción norteña del Golfo en el Plioceno Tardío, y 3) un canal marino que separó las poblaciones más meridionales cercanas al istmo de La Paz durante el Plioceno. El estudio de la tectónica brinda el marco de referencia para los grandes fenómenos geológicos, en este caso la deriva de la península y el ensamblaje histórico de sus componentes, mientras que el estudio de la distribución de las especies y su bagaje de información histórica contenida en su genoma ofrece la oportunidad de investigar fenómenos finos de la evolución geológica que no dejaron huellas claras ni en la tectónica ni en la columna estratigráfica.

Figura 3. A) Iguana de San Pedro Nolasco (Ctenosaura nolascensis). Especie endémica de la isla del mismo nombre. B) Vegetación típica de la región de Cataviñá en el área natural protegida “Valle de los Cirios” en Baja California. Izquierda, cirio (Fouquieria columnaris), centro, copalquín (Pachycormus discolor), derecha, cardón sagüeso
(Pachycereus pringlei). Al frente, Agave cerulata.
Conclusiones
La geodinámica descrita por la tectónica de placas modifica la repartición de los continentes y de los océanos. En el noroeste de México, el cambio de los movimientos relativos entre las placas tectónicas involucradas en esta parte del mundo ha desencadenado una ruptura continental y la deriva paulatina de la península de Baja California a partir del fin del Mioceno Medio (aproximadamente 12 Ma) y una transgresión marina progresiva en el Golfo de California en formación durante los últimos 10 Ma. Nació un océano en el sentido geológico de la palabra, y un mar en el sentido geográfico, en donde especies marinas provenientes del Pacífico ecuatorial encontraron oportunidades de establecimiento y de radiación adaptativa. Los nuevos ecosistemas son el resultado de los cambios tectónicos, oceanográficos y geomorfológicos. Entre éstos se cuenta el aumento de la profundidad, el vulcanismo submarino, cambios en la temperatura del agua, las surgencias, cambios en el nivel del mar, la alteración de corrientes superficiales y profundas, y los aportes detríticos desde el continente. El desarrollo de la tectónica de placas brindó un marco explicativo a la deriva continental de Wegener que describía la coincidencia de las costas y la similaridad biológica entre los continentes. Ahora, las técnicas moleculares permiten encontrar en el material genético de los organismos la firma de antiguos fenómenos de diversificación y aislamiento biológico, indicativos de eventos geológicos y tectónicos muy finos, que de otra manera no serían evidentes. El Golfo de California no tiene el futuro de los grandes océanos como el Atlántico que se abrió y continúa creciendo entre grandes continentes previamente unidos en el supercontinente Pangea, pero la naturaleza oceánica de la corteza de sus cuencas profundas indica la formación de un océano marginal provocado por el desprendimiento de un bloque continental. De igual manera, la separación de la Península y de las islas del Golfo provocó un cambio en las condiciones climáticas, hidrográficas y oceanográficas que influyeron de manera determinante en la fascinante evolución biológica de la cuenca del Golfo de California.
Agradecimientos
Este trabajo se realizó durante las estancias sabáticas de AB y AMY en la Universidad de Arizona, EUA, con ayuda del Programa de Apoyos para la Superación del Personal Académico-DGAPA, Universidad Nacional Autónoma de México. Los autores agradecen al M.C. Héctor Mendivil-Quijada por su apoyo en la elaboración de la figura 1 y a Enriquena Bustamente Ortega por su apoyo técnico.
*Instituto de Geología, **Instituto de Ecología
Universidad Nacional Autónoma de México, Hermosillo, Son.
Contacto: tcalmus@unam.mx
Referencias
Antonelis, K. (1999). GPS determination of current Pacific-North American plate motion, Geology, v. 27, p. 299-302.
Arreguín-Sánchez, F., Arcos H., E. (2011). La pesca en México: estado de la explotación y uso de los ecosistemas. Hidrobiológica 21(3): 431-462.
Brusca, R.C., Hendrickx, M.E. (2010). Invertebrate Biodiversity and Conservation in the Gulf of California In: Brusca, R.C. (ed.). The Gulf of California: Biodiversity and Conservation. Arizona-Sonora Desert Museum Studies in Natural History. University of Arizona Press, Tucson, Cap. 4, p.72-95.
Carmona, R., et al. (2015). Importancia del noroeste de México para la migración e hibernación de las aves playeras. Conabio. Biodiversitas, 121:6-11.
DeMets, C. (1995). A reappraisal of seafloor spreading lineations in the Gulf of California: Implications for the transfer of Baja California to the Pacific plate and estimates of Pacific-North America motion, Geophysical Research Letters, v. 22, p. 3345-3548.
Dixon, T., et al. (2000). New kinematic models for Pacific-North America motion from 3 Ma to Present II: Evidence for a “Baja California shear zone, Geophysical Research Letters, v. 27, p. 3961-3964.
Francheteau, J., et al. (1984). Nacimiento de un océano. Consejo Nacional de Ciencia y Tecnología, México, D.F., 92 pp.
Felger, R.S., Wilder, B.T., Romero-Morales H. (2013). Plant Life of a Desert Archipelago: Flora of the Sonoran Islands in the Gulf of California. University of Arizona Press, Tucson.
Hastings, P.A., Findley, L.T., van der Heiden, A.M. (2010). Fishes of the Gulf of California, In: Brusca, R.C. (ed.). The Gulf of California: Biodiversity and Conservation. Arizona-Sonora Desert Museum Studies in Natural History. University of Arizona Press, Tucson, Cap.5, p. 96-118.
Helenes, J., Carreño, A.L. (1999). Neogene sedimentary evolution of Baja California in relation to regional tectonics, Journal of South American Earth Sciences, v. 12, p. 589-605.
Holt, J.W., Holt, E. W., Stock, J.M. (2000). An age constraint on Gulf of California rifting from the Santa Rosalía basin, Baja California Sur, México, Geological Society of America Bulletin, v. 112, p. 540-549.
Luque, D., et al. (2016). Complejos bioculturales de Sonora: pueblos y territorios indígenas. Centro de Investigación en Alimentación y Desarrollo, A.C. Hermosillo, México.
Márquez M., R. (1990). FAO species catalogue. Vol. 11: Sea turtles of the world. An annotated and illustrated catalogue of sea turtle species known to date. FAO Fisheries Synopsis No. 125, Vol. 11. Rome, FAO. 81 p.
Ochoa-Landín, L., et al. (2000). Sedimentology and stratigraphy of the Upper Miocene Boleo Formation, and its relation with the opening of the Gulf of California, Santa Rosalia, Baja California, Mexico, Revista Mexicana de Ciencias Geológicas, v. 17, p. 83-95.
Ortlieb, L., Colletta, B. (1984). Síntesis cronoestratigráfica sobre el Neógeno y el Cuaternario marino de la cuenca de Santa Rosalía, Baja California Sur, México, In: Malpica-Cruz, V., et al. (eds.), Neotectonics and sea level variations in the Gulf of California area, a symposium: México, D. F., Universidad Nacional Autónoma de México, Instituto de Geología, Contributions volume, p. 241-260.
Oskin, M., Stock, J., Martín-Barajas, A. (2001). Rapid localization of Pacific-North America plate motion in the Gulf of California, Geology, v. 29, p. 459-462.
Oskin, M., Stock, J. (2003). Pacific-North America plate motion and opening of the Upper Delfín basin, northern Gulf of California, Mexico, Geological Society of America Bulletin, v. 115, p. 1173-1190.
Riddle, B.R., et al. (2000). Cryptic vicariance in the historical assembly of a Baja California Peninsular Desert biota. Proc. Nat. Acad. Sci. 97, p. 14438–14443.
Seminoff, J.A. (2010). Sea Turtles of the Gulf of California: Biology, Culture, and Conservation, In: Brusca, R.C. (ed.). The Gulf of California: Biodiversity and Conservation. Arizona-Sonora Desert Museum Studies in Natural History. University of Arizona Press, Tucson, Cap. 7, p.135-167.
Urbán, J. (2010). Marine Mammals of the Gulf of California: An Overview of Diversity and Conservation Status, In: Brusca, R.C. (ed.). The Gulf of California: Biodiversity and Conservation. Arizona-Sonora Desert Museum Studies in Natural History. University of Arizona Press, Tucson, Cap. 9, p. 188-209.
Velarde, E., y Anderson, D.W. (1993). Conservation and management of seabird islands in the Gulf of California: setbacks and successes, In: J. Burger, M. Gochfeld y D. Nettleship (eds.) Seabirds on Islands: threats, case studies and action plans. ICBP Technical Publication. Cambridge, p. 721-765.